
گونه و گونه زایی
این صفحه مربوط به موضوعات تحقیق دانشجویان کارشناسی ارشد دانشگاه شهید بهشتی برای درس گونه و گونه زایی است. هدف از قرار دادن این مطالب در اینجا کمک به علاقه مندان این درس میباشد. بدیهی است که مطالب بدون اشکال نیست. لطفا با ارسال ایمیل ما را در رفع این اشکالات یاری فرمایید.
لیست اسامی دانشجویان عزیز و موضوع مربوط به هر یک از آنها در انتهای صفحه قرار دارد.
Founder
Effects and Bottlenecks
Postzygotic
or Hybrid barriers
Synapomorphy
& Symplesiomorphies
-گونهزايي
طبقهبندي
مدلهاي
بالقوه گونهزايي
در موجودات
زندة داراي
توليد مثل
جنسي
By geography and level (after Mayr ,
1963)
1- Hybridization
(حفظ
هيبريدهاي
بين دو گونه
كه در لحاظ
توليد مثلي در
اين دو گونه
مجزا هستند.
2- Instantaneous
speciation يا گونهزايي
آني (از طريق
افراد)
-A Genetically
(يك موتاسيون)،
Cytologically -B
-a
آرايش مجدد
كروموزومها
-b
پليپلوئيدي
3- Gradual
speciation (از طريق
جمعيتها)
-A Sympatric speciation
-B ParaPatric speciation
-C Auopatric speciation (يا
جغرافيايي)
-a PeriPatric sPeciation (از طريق
تكامل يك كلني
مجزا)
-b vicariant speciation
(تقسيم محدودة
پراكندگي در
اثر يك مانع
دروني يا
انفراض
جمعيتهاي
مداخله كننده)
By PoPatation genetic mode (after TemPleton
, 1982)
1- Transilience
-A Hybrids maintenance
(انتخاب در
سطح هيبريدها)
-B Hybrid recombination
(انتخاب
بازتركيبي كه
بعد از هيبريد
شدن اتفاق ميافتد)
-C Chromosomal
(تثبيت آرايش
مجدد
كروموزمها در
اثر رانش ژن و
انتخاب طبيعي)
-D Genetic
(واقعة موسس
در يكي كلني)
2- Divergence
-A Habitat
(انتخاب
متفاوت بدان
جدايي از طريق
ايجاد فاصله)
-B Clinal
(انتخاب در
سطح cline
همراه با
جدايي از طريق
فاصله)
-C Adaptive
(بوجود آمدن
يك مانع
بيروني يك divergent
microeclation)
Symbiont- hdueed incompatibility
در
بعضي از حشرات
عقيمي
هيبريدها در
نتيجة فاكتورهاي
سيتوپلاسمي
ايجاد ميشود
و چنانچه
والدين در
مصرف آنتيبيوتيكها
قرار گيرند،
داراي قدرت
زيستن خواهند
بود. اين
ناسازگاري به
دليل آلودگي
با نوعي
باكتري (wolbachia)
كه به داخل
سيتوپلاسم
وارد ميشوند
صورت ميگيرند.
هنگامي كه
زنبورهاي
انگل wasonia vitripennis
و giraulti با
يكديگر آميزش
كنند، فقط
افراد را
بوجود ميآورند.
البته اين
افراد هيبريد
نيستند چرا كه
اين حالت يعني
نمو تخم بارور
نشده در
زنبورها زياد
اتفاق ميافتد.
هنگامي كه
نرمهاي هر
گونه تحت تاثير
آنتيبيوتيك
قرار گرفته و
سپس با مادههاي
گونة ديگر
خفتگيري كنند
درصد بالايي
از زادهها
هيبريدهاي
مادة بارور
هستند.
آزمايشات ميكروسكوپي
و تحزيه و
تحليلهاي
ژنتيكي نشان
دادند كه در
آميزشهاي
ناسازگار
كروموزمهاي
والدين در
سلول تخم
بارور نشده در
مراحل اولية
مينوز بخوبي
متراكم نشده و
ظاهراً از بين
ميروند. اين
تخمها تنها
داراي يك
كروموزوم
مادري هستند
كه منجر به
ايجاد نرهاي
غير هيبريد ميشود.
نژادي از اين
زنبورها كه
داراي
كروموزمهاي graulti و
سيتوپلاسم
حاوي vitriPennis
هيبريدهاي
مادهاي را
بوجود ميآورند.
با اين حال
آميزش آنها با
نرهاي آلوده يعني
هم گونة خود
با باكتريهاي
متفاوت graulti
منجر به ايجاد
افراد هيبريد
نميشود.
بنابراين
ناسازگاري
بين N.graulti, N. vitriPennis
به دليل
ناسازگاري
بين باكتريهايي
است كه اين دو
گونه حمل ميكنند.
The Genetic Basis of Prezygotic isolation
موانع
پيش تخمي در
بسياري از
جانوران
بويژه در
آنهايي كه
عموماً افراد
نر به هنگام
فصل جفتگيري
سيگنال ايجاد
ميكنند و
علاوه بر اين
سيگنالها تظاهراتي
نيز در آنها
ديده ميشود
پيچيده است.
اساس ژنتيكي
تغييرات صفات
مورفولويكي
به هنگام
خفتگيري به
منشا به اساس
ژنتيكي خود
صفات است.
مثلاً
همانطور كه
قبلاً اشاره
شد اختلاف
شكلسربين دو
گونة heteroncura, D.silvestris
منشاء پليژنتيك
دانسته و
كروموزوم x
در اين ميان
نقش مهمي را
ايفا ميكند.
نتايج بدست
آمده نشان
دادند كه
جدايي توليد
مثلي بين
جمعيتها
معمولاً در
سطح بيش از دولوكس
صورت ميگيرد.
در جدول صفحه
قبل اشاره شده
است كه يكي از
مدلهاي گونهزدايي
آني گونهزادي
ژنتيكي و آنهم
از طريق يك
موتاسيسون
است ولي ما
اين را قبول
نداريم. چرا
كه اولاً ممكن
است اين موتاسيون
در بعي افراد
ايجاد شود يا
اينكه در يك
فروهتروزيگوت
بوجود آيد كه
معمولاً اين
موتاسيسون
توسط انتخاب
طبيعي حذف ميشود.
هيچ مثال مانع
كنندهاي نيز
در اين مورد
وجود ندارد
تكامل موانع
توليد مثلي از
طريق
جايگزيني چند
آكل گونهزايي
تدريجي
ناميده ميشود
كه Mayr آن
را به سه شكل
تقسيم نموده
است.
- Allopatric
speciation
گونهزايي
آلوپاتريك يا
جغرافيايي
گونهزايي
آلوپاتيك
يعني تكامل
موانع توليد
مثلي ژنتيكي
بين دو جهت كه
از لحاظ
جغرافيايي
توسط يكي مانع
خارجي فيزيكي
مانند
توپرگرافي،
آب (يا زمين) يا
زيستگاههاي
نامساعد از
يكديگر جدا
شده باشند. در
گونههايي كه
انتشار كمي
داشته و يا
شديداً به
زيستگاه خود
محدود شدهاند
ممكن است
موانع خارجي
جمعيتها را در
مقياس
ميكروجغرافيايي
تقسيم كنند.
مانند زيستگاههاي
مجزايي كه در
يكي درياچه
وجود دارد.
آلوپاتري با
كاهش شديد
جابجايي افراد
يا گامتهاي
آنها مشخص ميشود
نه با فاصلة
جغرافايي و
موانع خارجي
بتاولات
ژنتيكي را
كاهش داده و
به صفر نميرساند.
دو نوع گونهزايي
آلوپاتريك
وجود دارد كه
از لحاظ
ساختار جمعيت
و ديناميك
ژنتيكي با
يكديگر
اختلاف دارند:
1) vicariont speciation يا
vicariance
زماني اتفاق
ميافتد كه در
جمعيت كه
داراي انتشار
وسيعي هستند
با ايجاد يك
مانع خارجي
انقراض
جمعيتهاي وارد
شده با مهاجرت
به يك ناحيه
مجزا تقسيم
شوند به عنوان
مثال ظهور
تنگة پاناما
در پليوسن بسياري
موجودات آبزي
را به دو قسمت
پاسيفيكي و
كارائيب
تقسيم نمود كه
بعضي از آنها
تبديل به گونههاي
مجزايي شدند.
بنظر ميرسد
كه اين نوع
گونهزايي با
انتخاب طبيعي
ورانش ژن در
دو جمعيت همراه
باشد و
احتملاً در
يكي از
جمعيتها
بيشتر از
ديگري است.
نوع ديگري از
گونهزدايي
آلوپاتريك
زماني اتفاق
ميافتد كه يك
كلني از يك
جمعيت مادري
كه داراي انتشار
وسيعي ميباشد.
اشتقاق پيدا
كند. Mayr
اين نوع گونهزايي
را Peripatric speciation
ناميده است.
بسياري از
متخصصان
ژنتيك جمعيت
معتقدند كه تغييرات
ژنتيكي ايجاد
شده در گونهزايي
vicariant, peripatric
شبيه هم ميباشند.
اما Templeton , Carson , Mayr
اظهار داشتند
كه به هنگام
تشكيل كلني و
تا زماني كه
اندازة جمعيت
آن پايين است
در اثر رانش
ژن تغييراتي
در فركانس
آللي كلني در
بعضي لوكوسها
ايجاد ميشود.
اين تغييرات
منجر ميشوند
تا انتخاب
طبيعي روي
لوكوسهاي
ديگر يا تركيبات
ژنتيكي كه
جديداً با
محيط سازگار
شدهاند عمل
كند. در اين
حالت جمعيت
يكي اوج تغيير
يا Peak shift را
متحمل ميشود
كه Templeton در
سال 1980 آن را تحت
عنوان Transilience
ناميده است.
اين مدل كه
تحت عنوان
گونهزايي
اثر مؤسس (founder
effect) نيز
ناميده ميشود
بسيار قابل
بحث است.
-
شواهد گونهزايي
آلوپاتريك
GeograPhic patterns
به
اين مطلب
اشاره شد كه
جمعيتهاي
جغرافيايي بيشتر
گونهها از
لحاظ ژنتيكي
با يكديگر
متفاوت ميباشند.
در بسياري از
موارد اين
تغييرات
سازشي بوده و
به دنبال اثر
انتخاب طبيعي
روي صفات مختلف
در محيطهاي
متفاوت ايجاد
ميشوند.
هنگامي كه در
اث ايجاد
موانع خارجي
شارژ ژن بين
جمعيتها تا
اندازة زيادي
كاهش پيدا كند
اين اختلافات
بيشتر خواهند
شد. نكتة قابل
توجه اين است
كه هنگامي كه
جمعيتهاي
جغرافيايي يك
گونه در
آزمايشگاه
مورد بررسي
قرار گيرند از
لحاظ شدت
جدايي پيش
تخمي و پس
تخمي با
يكيدگر
متفاوت خواهند
بود به عنوان
مثال در
جمعيتهاي
مجزايي نوعي
سمندر به نام Desmoynathus ochroPhaeus
جدايي توليد
مثلي از مقدار
كم تا زياد
تغيير ميكند.
در مثال circular
overloP در نوعي
سمندر به نام Ensation ديديم كه
چگونه كاهش شارژ
ژن ميان
جمعيتها باعث
تكامل جدايي
توليد مثلي ميشود.
-
ارتباط بين
موانع موجود
در گذشته و
حال
-
ميزان تمايز
جمعيتهاي
الوپاتريك به
فراواني و
بزرگي موانع
توپوگرافي
بستگي دارد.
به عنوان مثال
كه در نواحي
كوهستاني
زندگي ميكنند
بسيار زياد
است چرا كه در
اين مناطق
سيستمهاي
رودخانهاي
باعث جدا شدن
جمعيتها از
يكديگر ميشوند.
مثال ديگري كه
در اين زمينه
وجود دارد در
مورد گونههاي
ساكن جزيره
است. گونههاي
ساكن جزاير از
لحاظ اكولوژي
و رفتار در مقايسه
با انواع
مشابه خود در
خشكي تنوع
بيشتري دارند.
مواردي
از تهاجم مضاعف
يا double invadion
شواهدي
بيشتري براي
گونهزايي
آلوپاتريك
گنجشك كوچك
است كه در
قارة استراليا
انتشار
فرواني داشته
و جمعيت تمايز
يافتهاي از
آن نيز در
جزيرة مجاور
استراليا كه
تا سماني
ناميده ميشود
يافت ميشود.
در اين جزيره
علاوه بر گونة
مزبور گونة A-ewingi نيز وجود
دارد. گونة
اخير بدون شك
در همان ابتداي
شكل گرفتن
جزيرة
تاسماني
منشاء گرفته و
گونه A.pusilla
در اثر تهاجمي
كه اخيراً
صورت گرفته
وارد جزيره
شده است. اين
تهاجم
چندگانه
توضيح ميدهد
كه چرا هر يك
از جزاير آرشيپلاگو
چندين گونه
نزديك به هم
را در خود جاي
دادهاند. در
حاليكه در
جزيرة دور
افتادهاي كه
وسعتي مشابه
دارد، تنها يك
گونه انتشار دارد.
نواحي
تماس (contact zonez)
بين اشكال
تمايز يافته
غالباً در
نتيجة گسترش
جمعيتهايي كه
در ابتدا
آلوپاتريک
بودهاند
ايجاد ميشوند.
Avist ,
Bermingham
آزمايشاتي بر
روي DNA
ميتوکندريايي
نمونههاي
متعلق به 2
گونه از ماهيهاي
رودخانهاي
جنوب شرق
آمريكا انجام
دادند. در هر
يك از 2 گونه كه
به سه خانواده
تعلق داشتند
توالي DNA
جمعيتهاي
شرقي و غربي
در دو clade
متفاوت قرار
گرفت. اين
مطالعات نشان
ميدهند كه
زماني شارش ژن
ميان جمعيتهاي
شرقي و غربي
براي مدت
طولاني كاهش
يافته است و
اين دو جمعيت
در غرب
فلوريدا و شرق
آلاباما
يكديگ را
ملاقات ميكردهاند.
آنها
دريافتند كه
جدايي بين 3 تا 4
ميليون سال
پيش صورت
گرفته است. در
آن زمان به
دليل آب شدن
يخچال Pliocen interglacial
سطح آب به 0 تا 60 متر
بالاتر از سطح
كنوني رسيده و
بيشتر مناطق توسط
آب شور پوشيده
شدهاند و اين
امر باعث
ايجاد مانعي
براي عبور ماهيهاي
آب شيرين شده
است.
-
مطالعات
آزمايشگاهي
Genetic Modeis of Ailopatric sPeciation
براي
تكامل
ناسازگاري
بين دو جمعيت
سه مدل ژنتيك
جمعيتي را ميتوان
در نظر گرفت.
اين مدلها
عبارتند از:
گونهزايي
بوسيلة
انتخاب
طبيعي، گونهزايي
بوسيله رانش
ژن و گونهزايي
بوسيله Peak shift
كه تركيبي از
برانش ژن و
انتخاب طبيعي
است.
- Speciation
by natural selection
در
اين مورد در
بوانسكي و
مولر مدلي را
در سطح دولوكس
معرفي كردند
كه قابل تقسيم
به لوكوسهاي
متعد نيز ميباشد
در نظر بگيريد
كه ژنوتيپ
اجدادي در دو
جمعيت ![]() است.
تغييرات
ايجاد شده در
شرايط زيست
محيطي منجر ميشود
كه انتخاب
طبيعي در
جمعيت 1، A1
را به جاي A2
و B1 را
به جاي B2
برگزيند. ر
اين حالت
ژنوتيپ
جمعيتها
برابر خواهد
بود با به
ترتيب
است.
تغييرات
ايجاد شده در
شرايط زيست
محيطي منجر ميشود
كه انتخاب
طبيعي در
جمعيت 1، A1
را به جاي A2
و B1 را
به جاي B2
برگزيند. ر
اين حالت
ژنوتيپ
جمعيتها
برابر خواهد
بود با به
ترتيب![]() و
و ![]() برابر
با
برابر
با ![]() داراي
باروري و قدرت
زيست كمي شده
و هم جمعيتهاي
داراي
باروري و قدرت
زيست كمي شده
و هم جمعيتهاي
![]() از
طريق تفاوت در
رفتارهاي
جنسي يا ديگر
موانع پيش
تخمي از
يكديگر مجزا
ميشوند. در
اين مدل هيچيك
از جمعيتها از
مرحلة
هتروزيگوتي
عبور نميكند.
از
طريق تفاوت در
رفتارهاي
جنسي يا ديگر
موانع پيش
تخمي از
يكديگر مجزا
ميشوند. در
اين مدل هيچيك
از جمعيتها از
مرحلة
هتروزيگوتي
عبور نميكند.
ممكن
است در حالت
ديگري از اين
مدل جانشيني
پي در پي
آللها كه منجر
به جدايي
توليد مثلي ميگردد
در يكي از
جمعيتها رخ
دهد. در اين
حالت اگر
ژنوتيپ هر دو
جمعيت ![]() باشند
ممكن است در
يكي جمعيتهاي
حاصله
باشند
ممكن است در
يكي جمعيتهاي
حاصله ![]() و
و
![]() بوده
و از لحاظ
توليد مثلي
ناسازگار
هستند. طبق
اين مدل جدايي
اوليد مثلي
محصول جانبي
اختلافات
سازشي در دو
جمعيت ميباشند.
بوده
و از لحاظ
توليد مثلي
ناسازگار
هستند. طبق
اين مدل جدايي
اوليد مثلي
محصول جانبي
اختلافات
سازشي در دو
جمعيت ميباشند.
- Speciation
by genetic drift
در
اين حالت نيز
مدلي نظير مدل
فوق در نظر
گرفته ميشود
با اين تفاوت
كه در اينجا
رانش ژن به
جاي انتخاب
طبيعي باعث
جايگزيني
آللها ميشود
در اينجا نيز
واكنش متقابل
بين B1,A1
موجب ايجاد ناسازگاري
توليد مثلي ميشود.
- Speciation
by Peak Shift
اين
نوع گونهزايي
تحت عنوان
گونهزايي Peripatric يا گونهزايي
اثر مؤسس نيز
ناميده ميشود
اين حالت
زماني اتفاق
ميافتد كه
انتخاب طبيعي
و رانش ژن روي
يك جمعيت كوچكي
كه در تنگه
قرار گرفته
است اثر ميكنند
و زماني كه
كلني توسط
تعداد محدودي
از افراد تأيس
شود مشاهده ميشود
طبق اين مدل
در لوکوسهاي
شاخص رانش ژن
باعث بالا
رفتن فركانس
آلاي نادري ميشود
كه باعث پايين
آمدن Fitess
هتروزيگوتها
شدهاند. سپس
اين آللها
توسط انتخاب
طبيعي تثبيت ميشوند
اين عمل ممكن
است به نوبة
خود باعث
ايجاد انتخاب
اپيستاتيك در
فركانس آللي
لوكوسهاي
ديگر شود.
بدين ترتيب
كلني جديد از
لحاظ ساختار
ژنتيكي با
كلني اجدادي ناسازگار
خواهد بود نقش
انتخاب طبيعي
- Fcological
selection and SPeciation
در
آنجايي كه
اختلافات
موجود ميان
جمعيتهاي يك
گونه يا گونههاي
نزديك به هم
در نتيجه سازش
با شرايط
اكولوژيك
مختلف ايجاد
ميشود. بسياري
از زيست شناسان
جدايي توليد
مثلي را اثر
جانبي اختلاف
سازشي ميدانند
معمولاً
تغييرات
ژنتيكي كه
باعث جدايي پستخمي
ميشوند را به
زحمت ميتوان
به تغييرات
اكولوژيكي
نسبت داد ولي
نسبت دادن
تغييرات
ژنتيكي بوجود
آوردندة جداي
پيشتخمي
تغييرات
اكولوژيك
آسانتر است.
يي از محدود
مثالهاي
موجود
فينچهاي جزاير
گالاپاگوس
هستند. در اين
فينچها
تغيير اندازه
و شكل منقار
نوعي سازش با
رژيمهاي
غذاي متفاوت
محسوب ميشود.
اين فينچها از
لحاظ رنگ بدن
بسيار شبيه به
يكديگر ميباشند.
مثال
ديگري كه ميتوان
دربارة گونهزايي
توسط انتخاب
طبيعي با نام
علمي Gastorasteus
ميباشد جد
دريازي اين
ماهي كه در
غرب كانادا
زندگي ميكرده
است در چندين
رودخانه آب
شيرين موجب
پيدايش چندين
جمعيت شده است
كه اندازة
كوچكتري دارند.
علاوه
بر اين در چند
درياچه دو شكل
از اين ماهيها
با هم مشاهده
ميشوند. يكي
سري از آنها
كوچك وسطح و
عدهاي ديگر
بزرگ و كف زني
هستند كه از
همان جد دريازي
و طي دوبار
تشكيل كلني
متوالي ايجاد
شدهاند .
آزمايشات
نشان دادند كه
در هر بار
مقايسه مادههاي
بدن با توجه نوع
زيستگاه و در
اثر انتخاب
طبيعي ايجاد
شده است. چرا
كه ماهيها در
اثر تكامل
موازي در
مناطق مختلف
تكامل پيدا
كردهاند
بنابراين
انتخاب
اكولوژيك
باعث كونهزايي
مواربيشده
است.
- Sexual
selection and speciation
انتخاب
جنسي ممكن است
به صورت رقابت
افراد يكي جنس
(معمولاً
نرها) براي
بدست آوردن
جنس ديگر و يا ترجيح
(انتخاب) يك
جنس (معمولاً
ماده) به
خصوصيتي در
جنس ديگر باشد
ماده يك
خصوصيت را در
جنس ديگر
انتخاب ميكنند.
صفات
ثانويه جنسي
در نرها نقش
مهمي را در
اين دو حالت
ايفا ميكنند.
آزمايشات
نشان دادهاند
كه ترجيح ماده
در يك جمعيت
بستگي به
صفاتي دارد كه
نرها به هنگام
تظاهرات جنسي
از خود نشان
ميدهند. در
بيشتر مواقع
مادهها
نرهايي را
انتخاب ميكنند
كه صفات تحريك
كننده و
بيشتري داشته
باشند. رفتار
شناسان
معتقدند كه
نادهها به SuPernormal stimulas
پاسخ ميدهند.
اما در مورد
اينكه مادهها
چرا بايد چنين
ترجيحي داشته
باشند فرضيه هاي
متعددي وجود
دارد:
1-
ترجيح نتيجهاي
درياي فرايند
خود تقويت
كننده بنام Runway sexual Selectiow
است كه به كمك
آن ترجيح ماده
و صفات نر
يكديگر را
تقويت ميكنند.
2-
ترجيح يك
پديدة سازشي
است چرا كه
صفات نر با
صفاتي كه باعث
افزايش Fitness
فرزندان ميشوند
مرتبط ميباشند.
3-مادهها
صفاتي را
ترجيح ميدهد
كه توسط
فرزندان پسر
به درست رسيد
و به آنها كمك
ميكند تا
براي تعيف
قلمرو يا جنس
مخالف با
نرهاي ديگر
رقابت كنند.
4-
مادهها آن
دسته از صفاتي
را كه بلندتر،
روشنتر و يا
به عبارتي
مشخصتر از
ساير صفات در
محيط باشند را
راحتتر
تشخيص داده و
به اين طريق
ميتوانند
حفت خود را
سريعتر پيدا
كند.
5-
مادهها به
واسطة داشتن
اندامهاي حسي
و.يژه به سمت محركهاي
خاص تمايل
نشان ميدهند
اندامهاي حسي
خاص در ماده
در تكامل ترجيح
در مادهها و
نيز تكامل
صفات مختلف در
نرها نقش مهمي
را عهد داده
ميباشند.
معمولاً
جدايي Precygotic
به عنوان
مانعي براي
آميزش عمل ميكنند.
به عنوان مثال
چرا كه اولاً
صفات انتخابي
جنسي به عنوان
مانعي براي
آميزش عمل ميكنند.
به عنوان مثال
ويژگيهاي
صداي نوعي
قورباغه به
نام Physalaemus Pustulosus
با ترجيح ماده
انتخاب شدهاند.
مادههاي اين
گونه صداي
نرهاي خود را
به صداي نرهاي
ديگر گونهها
ترجيح ميدهند.
بنابرايت
اختلاف در
صداي ايجاد
شده به عنوان
يك مانع توليد
مثلي عمل ميكنند
دوماً
تاكسونهايي
كه انتخاب جنس
در آنها متنوع
و شديد است
داراي تعداد
گونههاي
زيادي هستند.
مانند
نوعهاي مگسخوار،
فروقاوليها،
اردكها و
نوعهاي بهشتي
و Drosophilaهاي هاوايي
در تمام اين
موارد بيشتر
گونهها از
لحاظ ژنتيكي
بهم شبيه ميباشند.
از اين رو
احتمال ميرود
كه اخيراً و
با سرعت منشاء
گرفته باشند در
تمام آنها
مادهها از
يكديگر تشخيص
داده نميشوند
اما نرها در
بسياري از
صفات با
يكديگر اختلاف
دارند.
- نقش
انتخاب طبيعي
در جدايي
توليد مثلي
تا
اينجا به اين
نتيجه رسيديم
كه جدايي
توليد مثلي
محصول جانبي
تغييرات
ژنتيكي ايجاد
شده توسط
انتخاب طبيعي
است. با اين
حال بسياري از
محققين
معتقدند كه
جدايي توليد
مثلي نوعي
سازش شده در
گونهها باعث
ميشود كه
احتمال آميزش
آنها با گونههاي
ديگر كمتر
شود. هنگامي
كه افراد جهش
نيافتند و A
با افراد B
آميزش پيدا ميكنند.
زادهها به
طور سازش نسبت
به pare specres در
وضعيت
بالاتري قرار
دارند. اما
زادههاي
افراد جهش
يافته نسبت به
زادههاي
افراد عادي در
وضعيت بهتري
قرار دارند.
در نتيجه
انتخاب طبيعي
باعث استقرار
و گسترش جهش
ميشود.
اما Mayr اظهار ميدارد
كه اگر چه
انتخاب طبيعي
ممكن است باعث
افزايش جدايي
توليد مثلي
شود اما موانع
توليد مثلي
غالباً محصول
جانبي اشتقاق
آلوپاتريك ميباشند.
Mayr
ايراداتي به
نظريه و بدين
قرار ميباشد. جدايي
جنسي در اشكال
آلوپاتريك
كامل وجود دارد
و در چندين
ناحية هيبريد
كه به نظر ميرسد
چندين هزار
سال عمر داشته
باشند، تكامل
پيدا نكرده
است؛ صفاتي كه
باعث پيشبرد جدايي
جنسي بين گونهها
ميشوند به
مناطقي كه
آنها در آنجا Sympatric بوده و با
خطر هيبريد
شدن روبرو
هستند محدود نميشود.
تصور
كنيد دو جمعيت
كه در اثر
گونهزايي
آلوپاتريك
بوجود آمدهاند
گسترش يافته و
تشكيل كيك
ناحية هيبريد
را دهند كه در
آن هيبريدها Fitness كمتري
نسبت به
ژنونيپ
والدين دارند
در اين مرحله سه
احتمال وجود
دارد: 1- ممكن
است نواحي
هيبريد تا مدت
زمان نامعيني
وجود داشته
باشند. 2- ممكن است
در اثر ايجاد
و گسترش
موتاسيون
هيبريد Fitness
برسد.
3-موتاسيون
ممكن است
گسترش پيدا
كند اما همانطور
كه دوژانسكي
بدان اشاره
كرده بود ممكن
است باعث كاهش
فركانس هيبريداسيون
شود. اما چه
چيزي مشخص ميكند
كه كداميك از
اين سه حالت
پيش خواهد
آمد؟
چنانچه
فركانس
موتاسيوني كه Fitness هيبريد
را به وضع
مطلوبي ميرساند
افزايش پيدا
كندفايدة
ژنهايي كه
باهث تشديد
جدايي توليد
مثل ميشوند كم
شده و در
نهايت باعث
ادغام شبه
گونههاي به
يك گونه واحد
ميشود.
انتخابي كه
باعث افزايش Fitness هيبريد
شود يك نكته
را در بردارد
انتخاب طبيعي
نميتواند
باعث شدت
يافتن جدايي
پيست تخمي
جمعيتهايي كه
تشكيل هيبريد
ميدهند شود.
يك سري صفات
وجود دارند كه
باهث كاهش
ميزان آميزش
دو جمعيت ميشوند.
اختلاف اين
صفات هنگتمي
كه دو جمعيت sympatric باشند
بيشتر از
زماني است كه
دو جمعيت
آلوپاتريك
باشند اين
حالت را Repoductive
character isotatcon نانامده
ميشود يعني
هيبريد در اثر
انتخاب طبيعي
حذف ميشود تا
جدايي توليد
مثلي صورت بگيرد.اين
فرايند علت
وقوع reproductive character
disPlacemen در واقع
كاهش Fitness
هيبربد باعث
انتخاب براي
تقويت جدايي
پيش تخمي ميشود. مثالي
كه در اين
مورد ميتوان
زد شباهت دو
گونه بومي
ساكن جزاير به
نامهاي DrosoPhila
mauritina با Simulens
آفريقايي است
و بنظر ميرسد
كه اين دو
گونه از گونه
اخير مشتق شده
باشند. با اين
حال انشعاب و
گونهزايي
سريع به خودي
خود نميتواند
شاهدي براي
گونهزايي Peak shift باشد.
چراد كه
انشعاب جمعيت
هنگامي كه در bottleneck قرار
دارد ممكن است
تنها در اثر
انتخاب طبيعي
صورت گرفته و
نميتواند در
اثر رانش ژن
ايجاد شود.
دانشمندان نشان
دادهند كه
جدايي جنسي
بين گونههاي
بهم DrosoPhila غالباً
اما نه هميشه
نامتقارن است.
گونهزايي
بالا در Drosophila
ساكن جزاير
هاوايي به bottleneck
ايجاد شده بين
جزاير و داخل
جزايره طي
تشكيل كلني و
بعد از آن
رابط ميشود
يك bottleneck در
اندازة جمعيت
براي ايجاد Peak shift در تعداد
كمي از
لوكوسهاي
انتخاب شده
كافي است.
بنابراين
جمعيتهاي
اجدادي و مشتق
شده بايد از
لحاظ آلوريم و
ديگر
ماركرهاي
مولكولي
اختلافات
شديدي داشته
باشند. اما
گونههاي
نزديكي بهم Drsophila ساكن جزايرها
هتروزيگوسيتي
و فاصله
ژنتيكي پليين
دارند. اين
حالت در بساري
از جفت گونههاي
نزديكي بهم
ديده ميشود و
قويترين
شاهد در برابر
founder effect و
دليل مهمي
براي گونهزايي
مي باشد. با
اين حالت در
بعضي موارد
گونههاي
مشتق شده bottleneck
را در اندازة
جمعيت سپري ميكنند.
-
گونهزايي
بوسيله آرايش
مجدد
كروموزومي
در
فصل 15 ديديم كه
گونههاي
نزديكي بهم از
لحاظ آرايش
مجدد
كروموزومي با
هم اختلاف
دارند كه اين
ممكن است باعث
كاهش با روي
هيبريد شود و
بنابران باعث
جدايي پس تخمي
ميشود. با
اين حال ظهور aneuplioid به
اندازهاي كه
انتظار داريم
زياد نيست و
مشخص كردن ميزان
عدم باروري
هيبريد در
نتايج
اختلافات ساختماني
كروموزومها
يا واكنش
متقابل ژنها
دشوار است.
لازمه جايگزيني
rearrangement كه
باروري
هتروزيگوتها
را كم ميكنند
اثر رانش ژن
جمعيت
بنابراين
تحليل اندازة
جمعيت ميباشد.
بنابراين
گونهزايي در
نتيجه اين
مكانيسم شبيه
گونهزايي founder cffect
بنابه دلايل
فوق آرايشهاي
مجددي كه
باروري
هتروزيگوتهارا
آهسته پايين
ميآورند.
نسبت به آرايشهاي
مجددي كه
باروري
هتروزيگوتها
را سريعتر يا
زيادتر پايين
ميآورد به
احتمال
بالاتري
تثبيت ميشوند.
هر چه جمعيت
كوچكتر بوده و
از bottleneck
عبور كند، اثر
بر روي بارري
شديدتر خواهد
بود. در اثر
جايگزيني
موفق
كروموزومها
احتمال وقوع
جدايي ژنتيكي
بيشتر خواهد
بود و هنگامي
كه چند
كروموزوم
جايگزين شوند
باروري به
آهستگي كاهش
پيدا ميكند
در حاليكه
جايگزيني يكي
كروموزوم
منفرد باروري
را شديد كاهش
ميدهد. عقيميهاي
هيبريد كه در
اثر اختلافات
كروموزومي ايجاد
ميشوند در
گياهان بسيار
ديده ميشوند.
مثلاً در دو
گونه از جنس Clarkia كه
كروموزومهايشان
از لحاظ دو trarislation و يك inversion
با هم اختلاف
دارند
هيبريدها باروري
كمي دارند و
بنظر ميرسد
كه يكي از
آنها والد
ديگري باشد
دانشمندان
معتفدند كه
اين گونهها
در اثر catadtroPhic
sPeciation بوجود
آمدهاند. اين
گياهان قادر
به خود تمامي
هستند و اين
پديده
احتمالاً
باعث افزايش
جايگزيني كروموزومها
ميشود.
-
شواهد
آزمايشگاهي گونهزايي
founder effect
محققين
بسياري
جمعيتها را
مكررأ از bottleneck
عبور دادهاند
تا ببينند آيا
جدايي توليد
مثلي تكامل پيدا
ميكند يا خير
و آزمايشات
نشان دادند كه
در پارهاي
موارد جدايي
بين bottleneck
والدين و در
هدهاي بيشتر
بين bottleneck
وجود دارد.
همكارانش
نشان دادند كه
گونهاي از
زنجرهها به
نام EnchenoPa binotato
متشكل از گونة
sibling
سيمپاتيك است
كه هر كدام به
جنس متفاوتي
از ميزبان
گياهي محدود
شدهاند اين
گونهها از
لحاظ توليد
مثلي از هم
مجزا هستند نه
تنها به اين
دليل كه روي
ميزبانهاي
مختلف جفتگيري
ميكنند بلكه
به اين دليل
كه اگر كنار
هم قرار گيرند
از لحاظ جنسي
از هم جدا ميشوند.
علاوه بر اين
چون تخمهاي
آنها در
زمانهاي
متفاوتي
تفريع ميشود
بلوغ جنسي و
جفتگيري آنها
نيز در فصل
تابستان در
زمانهاي
متفاوتي
انجام مي
گيرد. اين جدايي
آللوكروميك
احتمالاً توسط
گياه ميزبان
ايجاد ميشود.
Wood
معتقد است كه
شارژ ژن بين
گونهها
ابتدا به دليل
اختلاف زماني
سيكل زندگي آنها
كاهش پيدا
كرده و اين
كاهش باعث
ايجاد اختلاق
ژنتيكي در
ترجيح خفت و
ترجيح ميبان
ميشود. اين
فرضيه با مدلهاي
گونهزايي كه
در بالا
بدانها اشاره
شد متفاوت است
از اين لحاظ
كه جدايي تولد
مثلي در اثر
اختلاف
ژنتيكي آغاز
نشده و توسط
يك مانع خارجي
كه عدم
همزماني سيكل
زندگي آنها
بوده و اكولوژيك
ميباشد
بوجود آمده
است. اين مانع
شبيه موانع
گونهزايي
آلوپاتريك
است. مثالي كه
به مدل habitat isolation
ناشي از ايجاد
تغييرات
ژنتيكي نزديك
است مگس Rhagoletis
Pomonella ميباشد.
-
مطالعات
آزمايشگاهي
براي
اين منظور
دانشمندان
آزمايشات
زيادي روي DrosoPhila
انجام داده و
آنها را در
مصرف انتخاب DisruPtive قرار
دارند تا
بوجود آمدن
جدايي پيش
تخمي را بررسي
كنند. آنها single variation , deuble- variation را
كه مطابق كه
معادل انتخاب
جنسي و انتخاب
زيستگاه
هستند را مورد
آزمايش قرار
دادند. اگر linteage diseqwlibria بين ژنها
براي دو صفت
وجود داشته
باشد انتخاب
جنسي يا جدايي
جنسي اتفاق ميافتد.
اما چون جلوي
اين واكنش در
اثر recombination گرفته
ميشود،
جدايي توليد
مثلي اتفاق
نميافتد ولي
در single-vartion
اثر مخالف recombination
پايين است و
لذا جدايي
توليد مثلي
موفق است. معمولاص
در جمعيتهاي
طبيعي سؤالي
كخ وجود دارد
اين است كه
چگونه ممكن
است در عين
جال كه
كشاورزان
ميان دو جمعيت
.جود دارد
تركيبات
ژنتيكي مختلف
كه باعث جدايي
توليد مثلي جمعيتهاي
از يكديگر ميشوند
بوجود آيند.
جواب اين سؤال
اين است كه ممكن
است شدت
انتخاب
تركيبات
ژنتيكي مختلف
از ميزان شارژ
ژن بيت=شتر
باشد. هنگامي
كه شارژژن بين
دو جمعيت
افزايش پيدا
ميكند. در
صورت وجود
انتخاب طبيعي
قوي cline
شيبدار
ايجاد ميشود
به عنوان مثال
يكي از چند
گياه علفي است
كه اخيراً
نيبت به تحمل
سنگين تكامل
پيدا كرده است.
تحت تأثير
فشار انتخاب
طبيعي شديد
براي تحمل
فلزات سنگين
چندين جمعيت
از جمعيتهاي
همشايهاي كه
اين تحمل را
ندارد مشتق
شدهاند.
جمعيتهاي
مشتق شده از
لحاظ زمان گلدهي
نيز با جمعيت
و در متفاوت
بوده و مكرراً
خود لقاحي
انجام ميدهند.
- Sympatric
speciation
چنانچه
در محدودة يك
جمعيت Panmictic
جمعيتي كه
آميزش تصادفي
دارد يك مانع
بيولوژيك در
برابر
تبادلات
ژنتيكي ايجاد
شود. اين نوع
گونهزايي sympatric ناميده
شده و در اين
حالت هيچگونه
جبابي مكاني
از گونه اوليه
اتفاق نميافتد.
يكي از مدلهاي
گونهزايي
سيمپاتريكي
گونهزايي در
گياهان به روش
پليپلوئيدي
است كه در آن
يك تغيير آني
باعث جدايي
توليد مثلي يك
پليپلوئيد
جديد از گونه
اجدادي ميشود.
در بيشتر
مدلهاي گونهزايي
سيمپاتريك
انتخاب طبيعي
گسلنده است.
بزرگترين
مانع در برابر
گونهزايي
سيمپاتريك
اثر آنتاگونیست
موجود بين
انتخاب طبيعي
و نوتركيبي
است. انتخاب
طبيعي باعث
افزايش
ارتباط بين
آللها شده و
نوتركيبي اين
ارتباط را از
بين ميبرد.
- Polyploidy
and Hybrid speciation
چند
پديده در
گياهان
مشاهده ميشود
كه داراي نقش
قابل در گونهزايي
ميباشند. اين
پديدة
عبارتند از
1-
توليد مثل غير
جنسي توليد
مثل غير جنسي
در گياهان به
دو ردش تكثير
رويشي و aapomixis
(پارمتوژنريا
تخمك لقاح
نيافتد) صورت
ميگيرد.
توليد مثل غير
جنسي مفهوم
بيولوژيك
گونهزايي رد
ميكند و قبول
ميكند اما
تغييرات
مورفولوژيك و
اكولوژيك ايجاد
شده در اثر
توليد مثل
جنسي بيشتر
استو در بهضي
از گياهان نيز
مانند (Hieracra)
هم توليد مثل
جنسي را ميتوان
در آنها
مشاهده نمود.
بعضي از
هيبريدهايي
كه از نظر
جنسي عقيم
هستند به
فراواني يافت
ميشوند چرا
كه از طريق
آپوميكسيسي
بوجود آمدهاند.
2- خود
لقاحي
گياهاني كه
شديداًخود
لقاحي انجام
ميدهند از
لحاظ توليد
مثلي از بقيه
مجزا هستند علاوه
بر اين در خود
لقاحي
تغييرات
ژنتيكي و كروموزومي
به اندازهاي
هستند كه fitness
ژنوتيپها را
پايين
بياورند.
3-
جدايي
اكولوژيك و
سازش اكوتيپها
معمولاً در
گياهان بيشتر
از جانوران
ديده ميشوند.
اكوتيپها
جمعيتهاي
بسيار نزديك
بهمي هستند كه
از لحاظ
مورفولوژي
فيريولوژي و
زيستگاه با
يكديگر
اختلاف داشته
و در اثر
انتخاب
گسلنده از هم جدا
شدهاند.
جدايي پيش تخمي
در آنها بسيار
ضعيف بوده و
گاهي اوقات به
آنها زيرگونه
هم گفته ميشود.
بعضيها نيز
آنها را به
عنوان گونه
نيز شناخته ميشوند.
4- Hybridization
گياهشناسان
معتقدند كه
هيبريدزايي
در ميان گونههاي
گياهي بسيار
نزديك به هم
مكرراًديده
ميشود. گاهي
اوقات
هيبريدهاي
بين گونهاي
به گونهاي
مجزا تبديل ميشوند.
5- Polyploidy
موجود پليپلوئيد
به موجودي
گفته ميشود
كه بيش از دو
سري كروموزوم
داشته باشد و
در گياهان
بيشتر از
جانوران وجود
دارند. در واقع
پليپلوئيدي
يكي از روشهاي
اصلي گونهزايي
در گياهان
محسوب ميشود.
در اكثر موارد
گونههاي پليپلوئيد
در اثر هيبريد
شدن اجدادي كه
از لحاظ ژنتيكي
با هم اختلاف
دارند بوجود
ميآيند.
هنگامي كه يك
هيبريد تبديل
به يك گونة جديد
ميشود
فيلوژني از
حالت دو شاخهاي
خارج شده و
حالت شبكهاي
پيدا ميكند.
- پليپلوئيدي
ماهيت
پليپلوئيدي – پليپلوئيدها
از لحاظ توليد
مثلي از ديگر
اجداد پليپلوئيد
خود مجزا بوده
و از اين رو به
عنوان گونههاي
بيولوژيك
مجزا محسوب ميشوند.
گونهزايي
توسط پليپلویيدي
دو مشاهده
دارد:
1)
تنها روش گونهزايي
آني است كه
توسط يكي
رويداد
ژنتيكي صورت ميگيرد.
2) تنها روش
گونهزايي
سيمپاتريك كه
شواهدي زيادي
براي تصديق آن
وجود دارد.
پليپلوئيدي
در جانوران
بسيار نادر
است. پليپلوئيدهايي
كه توليد مثل
جنسي دارند در
محدودي از
ماهيها و
قورباغهها
وجود دارند و
بيشتر آنها
پازمترژنتيك
هستند. مانند
ملخها
سمندرها و
سوسمارها پليپلوئيدهاي
طبيعي بين دو
حد اتوپليپلوئيدي
آلوپلي
پلوئيدي
محدود شدهاند.
اتوپليپلوئيدها
در اثر ادغام
گامتهايي از
نظر ژنتيكي و
كروموزمي
شبيه به هم
بوده و به نظر
ميرسد متعلق
به يك گونه
باشند بوجود
ميآيند. به
عنوان مثال
سيبزميني
پرورشي (solanum
tuberosum)
اتوتتراپلوئيد
يك گونة
ديپلوئيد
آمريكاي
جنوبي است.
اما پلي
پلوئيدها از
ادغام
گامتهاي
افراد متعلق به
دو گونه بوجود
ميآيند.
اتوپليپلوئيدها
غالباً
باروري كمي
دارند. در
حاليكه در پليپلوئيدها
باروري به
صورت طبيعي
انجام ميگيرد
(علت اين امر
نحوة جدا شدن
كروموزمها در تقسيم
ميوز است كه
قبلاً درباره
آن توضيح داده
ده است) اما
حالتهاي حد
واسط بسياري
نيز ميتواند
وجود داشته
باشد.
-
گونه زايي در
اثر پليپلوئيدي
معمولاً
گياهي كه
پلوئيدي عجيب
(oddplody)
دارند (مانند
تريپلوئيدها
و
نيتاپلوئيديها)
تقريباً عقيم
هستند. چرا كه
قبلاً در يك
تريپلوئيد
هنگام تقسيم
ميوز و جدا
شدن جفت كروموزمها
در يك طرف دو
كروموزوم و در
طرف ديگر يك
كروموزوم
قرار ميگيرد
و چون منجر به
ايجاد
گامتهاي
آنئوپلوئيدي
ميشود باعث
عقيمي ميگردد
البته در
طبيعت پليپلوئيدهاي
عجيبي وجود
دارند كه به
روش غير جنسي
توليد مثل ميكنند
مانند جنس Cerpis
از Asterncea كه
داراي
جمعيتهاي 38، 58، 78
هستند) يك فرد
تتراپلوئيد
هم عقيم است
چون وقتي با
حد پلوئيد خود
آميزش پيدا ميكند،
هيبريد
تريپلوئيد
عقيمي را
بوجود ميآورد.
اين حالت براي
سطوح بالاتر
پلوئيدي هم وجود
دارد.
-
انتشار پلي
پلوئيدي
گياهشنسان
معتقدند كه
پليپلوئيدي
بيش از اتوپليپلوئيدي
منجر به ايجاد
گونههاي پليپلوئيدي
در گياهان ميشود.
چرا كه اتوپليپلوئيدها
باروري كمتري
دارند. در
بساري از جنسها
هيبريداسيون
منجر به ايجاد
پليپلوئيدهايي
ميشود كه از
لحاظ تعداد
كروموزومها
متنوع ميباشند
و يا منجر به
ايجاد
كمپلكسهاي
پليپلوئيد
ميشود. در
بسياري از جنسها
پليپلوئيدها
در مقايسه با
منسبوين و
پليوئيد در عرضهاي
جغرافيايي
بالاتر و در
محيطهاي
استرسزا
ويژه نواحي يخ
زده ميشوند.
بسياري از
گياهان پليپلوئيد
در اجرا پليپلوئيد
بوجود آمدهاند
در حاليكه عدهاي
از پليپلوئيدها
از اجدادي كه
تعداد
كروموزمهاي
كمتري داشتهاند
بوجود آمدهاند.
تكثير
و قدرت زيست
پلي
پلوئيدهاي كه
جديداً بوجود
ميآيند به
چند عامل
بستگي دارد.
خود لقاحي
داشتن Fitness
بالاتر از
پلوئيدها و يا
جدايي منبع
آنها از منابع
ديپليوئيدها
(يعني اشتغال
نمودن
زيستگاههاي
متفاوت)
بسياري از
تاكسونهاي
پليپلوئيد
از تاكسونهاي
اجداد خود در
زيستگاه و پراكندگي
متفاوت ميباشند
به عنوان مثال
در غرب آفريقا
علف دپيلوئيد
در زيستگاههاي
مرطوب
تتراپلوئيد
در زيستكاههاي
خشك (در پاي
تپههاي شني)
و هگزاپلوئيد
در نواحي
بسيار خشك و
شنهاي داغ
زندگي ميكنند.
اما مشخص نيست
كه اين
اختلافنا چه
حد ناشي از
پلي پلوئيدي و
تا چه اندازه
به دليل تغييرات
تكاملي است.
شاهد نشان ميدهند
كه پليپلوئيدي
در اين ميان
اهميت زيادي
دارد. افزايش
پلوئيدي باعث
افزايش
اندازة سلول و
ميزان آب آن
نمو آهستهتر
اندازة بزرگ
بدن و غالباً
بالا بودن سطح
بعضي از آنزيمها
هورمونها و
ساير تركيبات
شده و گياه را
در برابر حملة
گياهخوران و
پاتوژنها
محافظت ميكنند.
بنابراين
بسياري از پليپلوئيدها
به محض بوجود
آمدن
قادرنداكولوژيك
جديدي را
اشغال نمايند.
پليپلوئيدي
ممكن است باعث
ايجاد
قابليتهاي
فيزيولوژيك و
اكولوژيك
جديدي شود اما
باعث ايجاد
تغيير در
خصوصيات
مورفولوژيك
همچون تغيير در
ساختمان كل يا
ميوه نميشود
بنابراين پليپلوئيدي
باعث بوجود
آمدن يك جنس
جديد يا يك تاكسون
بالا درگير
نميشود.
-
بوجود آمدت
گونهها در
اثر
هيبريدشدن
هيبريديزاسيون
گاهي اوقات
منجر به گونههاي
مجزايي ميشود
كه از لحاظ
پلوتيدي شبيه
والدين خود ميباشد
Grant در
سال 1971 اين نوع
گونه زايي را Recombinational Specration
ناميد اين
پديده در
گياهان حداقل
در شرايط آزمايشگاهي
ديده شده است
اما در
جانوران به
ندرت ديده ميشود.
گونههاي
هيبريد
ديپلوئيد
معمولاً
داراي خصوصيات
مورفولوژيك
مولكولي و
كروموزومي
خاصي هستند.
-
سرعت گونهزايي
سرعت
گونهزايي
نظير سرعت
پيدايش
هرتاكسون
بالاتري به دو
مفهوم مرتبط
برميگردد:
سرعت
تكامل جدايي
توليد مثلي در
جمعيتهاي
متعلق به يك
گونه و تعداد
گونههايي كه
در واحد زمان
از گونه روي
بوجود ميآيند.
البته مورد
اخير معمولاً
برميگردد به
ميزان انشعاب
كلا و سرعت
افزايش تعداد
گونهها در يك
گروه
مونوفيلتيك
به سرعت
پيدايش و انقراض
آنها بستگي
دارد. مدت
زمان لازم
براي تكامل جدايي
توليد مثلي
نيز بسيار
متفاوت است.
مثلاً شناسي و
مولكولي حاكي
از آنند كه
مدت زمان لازم
براي جدايي
كامل توليد
مثلي حدود 3
ميليون سال
است. اما با
وجود اين
جدايي طولاني
هنوز هم بعضي
از جمعيتها از
لحاظ توليد
مثلي با هم سازگار
بود و
هيبريدهاي
باروري را
بوجود ميآورند.
بعضي از عوامل
باعث تسريع
گونهزايي ميشوند.
به عنوان مثال
در DrosoPhila
جزاير هاويي
ميزان انتشار
كم وجود فرصتهاي
بسيار براي
جدا شدن
جمعيتها از
طريق تشكيل
كلني به سرعت
انتخاب جنسي و
تنوع سريع
صفات جنسي
باعث تسريع
گونهزايي ميشوند.
علاوه بر فرصتهاي
اكولوژيكي يك
سري خصوصيات
دروني موجود زنده
نيز ممكن است
روي سرعت گونهزايي
تاثير
بگذارند. در
اين مورد ميتوان
مثال زد در
قورباغهها
اندام شنوايي
داخلي ميتواند
كوتاه تا بلند
باشند اندامهاي
شنوايي كوتاه
كه نسبت به
طيف باريكي از
موكانسهاي
پايين حساسند
در قورباغهها
اوليه و اندامهاي
شنوايي بلند
كه نسبت به
طيف وسيعي از
فركانسهاي
بالا حساسند
در قورباغهها
پيشرفته و
آنهايي كه
اخيراً منشاء
گرفتهاند
وجود دارند.
در دما نهايي
كه اندامهاي
شنوايي بلندي
دارند بسيار
متنوعتر ميباشد.
Ryan در
سال 1990 اظهار
داشت كه اندامهاي
شنوايي بلند
در مادههايي
كه واجد آن ميباشند
كمك ميكند تا
بتوانند صداي
نرها را بهتر
شناسايي كرده
و جدايي جنسي
و متقاباً
گونهزايي با
موفقعيت
بيشتري انجام
پذيرد بنابراين
فاكتورها
مؤثر بر گونهزايي
بدين شرح
خلاصه ميشوند:
1-
بوجود آمدن
فرصتهاي
مناسب براي
جدا شدن
جمعيتها كه
معمولاً بوسيلة
موانع
توپوگرافيك
يا ديگر موانع
ايجاد ميشود.
2-
پايين بودن
ميزان مهاجرت
و كاهش شارژ
ژن ميان
جمعيتها
3-
اختصاصي بودن
شرايط
اكولوژيك كه
اين امر موجب
محدود شدن
گونهها به
زيستگاهها
4-
انتخاب جنسي
قوي كه منجر
به ايجاد تنوع
در سیستمهاي
تشخيص جفت ميشود/
5- به
تنگنا رسيدن (bottleneck) اندازه
جمعيت كه باعث
تسهيل Pcak shift
ژنتيكي و
جايگزيني
آرايش
كروموزومي ميشود.
6-
اختلاف در
سيستمهاي
ژنتيكي و نمو (CPistatic Jenesystem)
Allopatry ,
Sympatry , Parapatry , Peripatry
گونه
زایی فرایندی
تکاملی است که
در تمام طول
دوران ها از
بدو پیدایش
اولین
موجودات زنده
روی زمین که
سلول های
پروکاریوتی
بوده اند تا
زمان حال در میان
تمامی
موجودات زنده
بطور آهسته و
پیوسته در جریان
بوده است و
همچنان ادامه
دارد و بطور
خلاصه می توان
گفت گونه زایی
از آنجا ناشی
می شود که در
جمعیتی از
اعضای یک گونه
آن چنان تفاوت
های کلیدی در یکسری
خصوصیا ت ژنتیکی
، رفتاری ، زیستگاهی
و ..... ایجاد می
شود که اعضای
این جمعیت دیگر
قادر به آ میزش
وتولید نسل طبیعی
با اعضای گونه
مبدا خود
نخواهند بود و
در حقیقت گونه
جدیدی از گونه
مبدا انشعاب می
یابد.
در
مبحث مکانیزم
های مربوط به
گونه زایی ،
بر اساس قلمرو
جغرافیایی که
جمعیت های در
حال گونه زایی
در آن از
یکدیگر
جدا می شوند
چهار طریقه در
طبیعت بطور
ملموس شناخته
شده است :
1-Allopatric 2- Sympartri 3-Parapatric 4- Peripatric

Allopatric Speciation
لغت
آلوپاتری
مشتق از دو
کلمه یونان
باستان است:
Allo به
معنای other ( دیگر) و patra به معنای father
land (سرزمین
پدری ، وطن)
.آلوپاتری
وآلوپاتریک
در مورد گروه های
موجودات زنده
ای بکار می
رود که اگرچه
خویشاوند بسیار
نزدیک هم
باشند در
قلمرو جغرافیایی
کاملا
جداگانه ای
زندگی می کنند
. گونه زایی
آلوپاتریک که
گونه زایی
جغرافیایی نیز
نام می گیرد
پدیده ای است
که به موجب آن
به واسطه یک
سد فیزیکی
خارجی مثل پدیده
کوه زایی ،
اشتقاق قاره
ها ، ایجاد یک
گسل بزرگ زمین
شناسی و....... در
درون جمعیت هایی
که از اعضای یک
گونه مبدا
هستند شکاف و
فاصله ای بزرگ
ایجاد شده و
جریان ژنی gene
flow ) ) بین دو
جمعیت قطع می
شود و طی نسل
های متمادی ،
هر جمعیت جدا
شده بطور
مستقل دچار تغییرات
ژنتیکی طبیعی
خاص خود شده و
در معرض فشار
گزینش طبیعت
قرار می گیرد .
نتیجه
این
است که به
مرور زمان طوری
این دو جمعیت
دچار واگرایی
ژنوتیپی و یا
فنوتیپی می
شوند و طوری
متحول می شوند
که اگر بعد از این مدت سد بین
آنها شکسته
شود و از میان
برود و اعضای
آنها با هم
تماس پیدا
کنند باز هم
افراد آنها
قادر به آمیزش
و تولیدمثل با
افراد گروه
مقابل
نخواهند بود.
چراکه عوامل
درونی ، جدایی
فیزیکی را تکمیل
کرده اند وآن
دو جمعیت
اکنون به دو
گونه متمایز
مبدل شده اند.


Sympatric Speciation
لغت
سمپاتری مشتق
از دو کلمه یونان
باستان است :
1-ریشه
sym به
معنای same ( مشابه ) ، like ( شبیه ) ، similar (یکسان) و .......
2-کلمه
patra به
معنای وطن ،
سرزمین پدری
گونه
زایی سمپاتریک
در واقع یک
واگرایی ژنتیکی
است که از یک
گونه والد
منفرد ، جمعیت
هایی گوناگون
حاصل می آیند ,در حالیکه
این جمعیت های
متفاوت حاصل از یک گونه ، در منطقه جغرافیایی
یکسانی زیستگاه دارند .
اشتقاق
گونه ها از
نوع سمپاتریک
فقط تحت تاثیر
عوامل درونی
(ژنتیکی) است و این شکاف و
جدایی بین جمعیت
های یک گونه می
تواند در ویژگی
های چند ژنی نیز
رخ دهد.اشتقاق
سمپاتریک می
تواند از
رقابت ها و
کشمکش های جنسی نیز نتیجه
شود که باز هم
به خصلت های ژنتیکی
متفاوت برمیگردد.گاهی
در روند
انتخاب طبیعی
هوموزیگوت بودن از هتروزیگوت
بودن مساعد
تر است
که
همین هم می
تواند به گونه
زایی سمپاتریک
منجر شود و این
مکانیزم به
نامDisruptive selection می باشد.



گونه
زایی سمپاتریک
در گیاهان بسیار
زیاد رخ می
دهد. سایر
مثال های آن نیز
در حشراتی است
که در منطقه ای
یکسان به میز
بان های گیاهی
متفاوتی
وابسته می
شوند . در هر
صورت باید گفت
در گونه زایی سمپاتریک
,گونه
ها در حالیکه
جایگاه مشابهی
را اشغال کرده
اند و در زیستگاه
یکسانی سکونت
دارند ، از
همدیگر واگرایی
و انشعاب پیدا
می کنند.

Parapatric Speciation
لغت para در
معانی beside (کنار) ، near (نزدیک) ، contrary (مقابل ،
معکوس ) و beyond (آن سوی ) بکار
می رود. در
گونه زایی
پارا پاتریک ،
قلمرو های دو
جمعیت واگرا
از هم جداست
ولی در برخی
نقاط هم پوشانی
دارند . در
واقع دو قلمرو
فقط یک جدایی
جغرافیایی
نسبی دارند
ودر کنار هم و
نزدیک هم بوده
و ساکنان این
دو قلمرو نیز
که خویشاوند
نزدیک یکدیگرند
هریک بطور
مستقل روند
تکاملی خود را
طی می کنند . ولی
هر چند وقت یکبار
برخی افراد از
هر گونه در
عرض مرز جغرافیایی
با افراد گونه
مقابل تماس بر
قرار می کنند
که گاهی ممکن
است به ایجاد Hybrid zone در مرز
ها بیانجامد.
گونه
زایی
پاراپاتریک
حاصل از
انتخاب طبیعی
است که به
دنبال
گوناگونی در
رفتارهای جفت یابی
جمعیت های
درون یک گونه
رخ داده و
مانع از جریان
ژنی شده و
باعث می شود
که این جمعیت
ها در امتداد یک
زیستگاه پیوسته و در مجاورت و
همسایگی با هم
هر یک به راه
خود روند.


Peripatric Speciation
لغت peri به
معنای حاشیه و
پیرامون می
باشد .
در
گونه زایی پری
پاتریک ، جمعیت
هایی کوچک
متعلق به یک
گونه واحد در
قلمروهای
جداگانه ای پیرامون
جمعیت اصلی ،
شکل می گیرند
که همین جدایی
از تبادل ژن
های آنها با
خزانه ژنی جمعیت
اصلی ممانعت می
کند و به
مرورعواملی
مثل Bottle neck و
Founder effect و
Genetic drift این
جدایی را که
در ابتدا فقط
حالت جغرافیایی کوچکی داشته
، تکمیل می
کنند.از معروف
ترین مثال های
گونه زایی پری
پاتریک در پشه
زیرزمینی شهر
لندن است که
گونه اصلی آن
در قرن نوزدهم
وارد زیر زمین
های لندن شده
و تا به امروز
گونه های
متعددی در
اطراف آن به
وجود آمده
اند.


Reference:
Wikipedia:speciation,allopatry,sympatry,parapatry,peripatry.
تكامل(سازگاري)شعاعي
يك
خصوصيت
تكاملي شعاعي
است كه به
وسيله ي افزايش
تنوع
مورفولوژيكي
و اكولوژيكي
از يك گونه به
سرعت گونه هاي
متنوعي بوجود
مي آيدودر اثر
ايجاد ويژگي
هاي جديد و
مفيد فنوتيپ
در پاسخ به
محيط سازش
ميابد.اين يك
فرايند
تكاملي مشتق
شده از انتخاب
طبيعي است.
تكامل
يك ساختار
جديد ممكن است
سبب شود يك
شاخه(تنوع)جديد
به وسيله پراكندگي
در نواحي
مختلف و قابل
دسترس بوجود بيايد.
يك مثال
كلاسيك از آن
تكامل چهارمين
نوك هلال در
دندان
پستانداران
است.اين ويژگي
سبب افزايش
تنوع مواد
غذايي مي شود
كه جاندار مي
تواند از آن
تغذيه كند و
تكامل اين
خصوصيت سبب مي
شود تعداد نيچ
هاي اكولوژيك
در دسترس كه
پستانداران
مي توانند
استفاده كنند
افزايش
يابد.اين
ويژگي موجب شد
كه كه از يك
گروه گروه هاي
مختلفي در طول
دوره
سنوزوييك بوجود
بيايد.واين
نمونه اي از
همان تكامل
شعاعي است.
پرندگان راه
ديگري را براي
ايجاد گروه
هاي مختلف
پيدا
كردند.مثلآ
توانايي پرواز
مسير جديدي
رابراي تكامل
اكتشاف كردن
آنها باز
كردواين آغاز
يك تكامل
شعاعي بود.
تكامل شعاعي
اغلب در نتيجه
پراكندگي يا
رشد يك ارگانيسم
دريك محيط با
نيچ بدون سكنه
به طور مثال در
يك درياچه
تازه شكل
گرفته يا يك
جزيره جدا شده
از قاره رخ مي
دهد. تكثير و
پراكندگي
جمعيت ممكن
است به سرعت
صورت بگيرد و
گونه مورد نظر
در سراسر نيچ
هاي ممكن
پراكنده
شود.در درياچه
ويكتوريا(يك
درياچه
ايزوله كه
اخير در دره Africanآشكل
گرفته)
به
طور شعاعي از
يك گونه يcichlidبيش
از300گونه ماهي
والديني و فقط
در مدت 15000 سال سازگاري
پيدا كرده اند
. تكامل شعاعي
عمومآ بعد از
يك انقراض بزرگ
اتفاق مي
افتد.چون به
دنبال يك
انقراض بسياري
از نيچ ها
خالي مي
شوند.به طور
مثال مي توان
از جايگزيني
دايناسور هاي
غير
پرنده(دايناسور
هايي كه قدرت
پرواز
نداشتند)با
پستانداران در
پايان دوره ي
كرتاسه و
جايگزيني بازوپايان
به وسيله ي
دوكفه اي ها
در حدود
ترياسه پيشين
اشاره كرد.
در شكل
زير نمونه اي
از تكامل
شعاعي را
مشاهده مي
كنيد.

1)گونه
A از قاره به
اولين جزيره
مهاجرت مي كند
2)در
اثر جدايي
قاره گونهAبه گونه B تبديل
مي شود
3)گونه
B به
جزيره ي دوم
مهاجرت مي كند
4)گونه
Bبه گونهC تبديل
مي شود.
5)گونه Bبه جزيره اول
مهاجرت مي كند
ولي ديگر قادر
به آميزش با
گونه C نيست.
6)گونه C به جزيره سوم
مهاجرت مي
كند.
7)گونه
Cبه گونه
D تبديل
مي شود.
8)گونه
D به
اولين و دومين
جزيره مهاجرت
مي كند.
9)گونه
D به گونهE تبديل
مي شود.
اين
فرايند مي
تواندبه طور
نامحدودپيش
برود تا جايي
كه به يك تنوع
گسترده برسد.
رشد
آلومتریک:
رشد
آلومتریک به ویژه
عصب بویایی در
دو گونه
خانواده Percidae
Stizostedion Lucioperca
Perca
Fluviatilis (Pisces)
در این
دو گونه رشد آلومتریک
در بعضی از
قسمت ها از
جمله encephalonسر
پوزه و بدن در
سنین مختلف
مشاهده می
شود.این رشد
آلو متریک
عبارت است از
ریتم کند در
رشد encephalon
رشد با ریتم
سریع تر در
قسمت های دیگر
که سریع ترین
ریتم در عصب
بویایی
مشاهده می
شود.
تنوع
خاص و زیاد
رشد در تلئوست
ها در ارتباط
است با شکل و
اندازه آنها و
متناسب است با
اجزای تشکیل
دهنده آن و هم
چنین موقعیت
بدنی و روش
زندگی آنها.
در این
مطالعه از
افرادی با
اندازه های
مختلف متعلق
به دو گونه
خانواده Percidae استفاده
کردیم.که
خانواده رایجی
در آب های
رومانی هستند.
21فرد
از Stizostedion Lucioperca
و12 فرد از Perca
Fluviatilis (Pisces) آنالیز
شده اند.هر
دوگونه شکارچی
هستند.
برای هر
فرد طول بدن (L) طول
سرLh) ) طول پوزه (Lm) طول( Len) encephalon وطول
عصب بویایی(Lofc.N) را
بررسی کردیم.
در
جدول یک طول این
قسمت ها لیست
شده است برای طول( Len) encephalon وطول
عصب بویایی(Lofc.N) دامنه
تنوع هم اضافه
شده است. .
از
آنجا که ریتم
رشد قبل و بعد
بلوغ جنسی
متفاوت است
وبرای هر گونه
با یک طول معین
بدن در ارتباط
است. به همین
دلیل ما گروه
های مختلف سنی
را برای طول
های مختلف
آنالیز کرده ایم
این مفادیر در
جدول دو لیست
شده ودر شکل یک
نشان داده شده
اند.
سفید
پوزه خاکستری
روشن سر سیاه encephalon خاکستری تیره عصب بویایی
.
در
مورد Stizostedion Lucioperca
برای افرادی
که اندازه
کوچکتری
دارند (201 -300 mm) رشد
عصب بویایی ریتم
سریع تری دارد(14%) سر و
پوزه بهطور
متوسط به میزان
7%
طویل شده اند. encephalon به
طور متوسط 4% طویل
شده اند. اما
رشد آهسته تر
در طول encephalon
با رشد سریع
تر عرض آن
جبران می شود.به ویژه
در یک گونه از tuber ها که
دارای مراکز بینایی
هستند که برای
گونه های
شکارچی ضروری
است.
نتایج
اندازه گیری mesencephal د ر گروه های سنی
مختلف
برای این
نمونه ریتم سریع
رشد عصب بویایی
مطابق با رشد
طول بدن است
از301 تا 350 میلیمتروقتی
طول عصب بویایی
تا 30% افزایش می یابد
encephalon سر و پوزه هم
طویل می شوند.
واین سریع ترین
ریتم رشد را
نشان می دهد.
واین هم زمان
با سه سالگی
است وقتی به
سن بلوغ رسیده
است.بعد از این
سن encephalon سر و
پوزه هم چنان
رشد می کنند
ولی ریتم رشد
بسیار آهسته
تر است
در
مورد Perca fluviatilis,که یک
گونه شکار چی
است سریع ترین
ریتم رشد برای
تمام تشکیلات
فوق وقتی است
که طول بدن از 100به 200 میلی
متر میرسددر این
دوره ریتم رشد
encephalon از
همه آهسته تر
است از آنجا
که هر دو
نمونه شکار چی
هستند بینایی
قوی برای آنها
بسیار ضروری
است به همین
دلیل تشکیلات encephalon بیشترین
تکوین را یافته
اند.هم چنین
درmesencephalon رشد
طولی بیشتر از
رشد عرضی است.
رسد
تمام بخشهای
مطالعه شده
آلومتریک است
سریع ترین ریتم
رشد در عصب بویایی
و آهسته ترین
در
encephalonمشاهده
می شود.
مقدمه
بر
مبنای تعریف بیولوژیکی
گونه اگر جریان
ژنی بین جمعیتهای
یک گونه توسط
تفاوتهای بیولوژیکی
که به عنوان
سدهای تولیدمثلی
شناخته می
شوند قطع شود،
گونه های
متفاوت از آن
جمعیتها به
وجود می آید.
اگر تبادل ژن
بین دو جمعیت
صورت بگیرد،
حتی اگر آن دو
از نظر
مورفولوژی خیلی
متفاوت باشند
از یک گونه
اند و برعکس
اگر جدایی تولید
مثلی بین دو
جمعیت وجود
داشته باشد حتی
اگر دو جمعیت
از نظر
مورفولوژیکی
از هم قابل
تمایز
نباشند، از دو
گونه مجزا
هستند. در نتیجه
گونه زایی از
تکامل سدهای بیولوژیکی
برای جریان ژنی
به وجود می آید
(Futuyma, 1998).
فاکتورهایی
که منجر به
جدایی تولید
مثلی می شوند
را می توان در
دو طبقه جای
داد:
1.
فاکتورهای پیش
زیگوتی که قبل
از اینکه لقاح
اتفاق بیفتد
وجود دارند.
2.
فاکتورهای
پس زیگوتی که پس
از لقاح اتفاق
می افتند.
از
جمله سدهای پیش
زیگوتی، جدایی
رفتاری است.
جدایی
رفتاری
جدایی
رفتاری شامل
تفاوتها در
رفتار طلب
جفت، سیگنالهای
شیمیایی،
صدا، رنگ و
مورفولوژی
است که به
افراد اجازه می
دهد تا اعضای
هم گونه خود
را بیابند. در
جانوران این
مقوله احتمالا
سدی مهم برای
جریان ژنی در
گونه های
هموطن (sympatric)
است. معمولا
در جانوران،
رفتار جفت یابی
(معمولا نرها)
به اعضای گونه
های دیگر نشان
داده نمی شود
و اگر هم این
کار انجام شود
این رفتار پذیرایی
برای جفتگیری
نخواهد داشت.
سیگنالها
و پاسخها
معمولا هر دو
اختصاصی
هستند، یعنی طی
گونه زایی از
هم واگرا شده
اند. به عنوان
مثال درجیرجیرکها
افراد گونه های
مختلف آوازهای
متفاوتی می
خوانند و ماده
ها فقط به
آوازهای
افراد هم گونه
خود پاسخ می
دهند (Futuyma, 1998). به طور
مشابه tarsiers
ساکن در یک
مکان جغرافیایی
مشترک صداهای
جفتگیری
متفاوتی
دارند.
معمولا
اکثر گونه های
جانوری از
ارتباط شیمیایی
استفاده می
کنند، این ترکیبات
شیمیایی به
کار برده شده
فرومونهای
جنسی خوانده می
شوند. برای
مثال پروانه
های نر به
غلظتهای خیلی
کم فرومونهای
جنسی جنس ماده
هم گونه پاسخ
می دهند. به
طوریکه وقتی
در تونلهای
بادی قرار
داده شدند،
پروانه های نر
تلاش می کنند
با یک شیءای
که به آن
فرومون جنسی
گونه خودش تلقیح
شده است، آمیزش
کنند اما به
فرومونهای سایر
گونه ها پاسخی
نمی دهند (Futuyma, 1998).
در
بسیاری از
حشرات،
پرندگان و ماهی
ها سیگنالهای
بینایی در
انتخاب جفت
فاکتوری مهم
است.در میان
پرنده ها:
اردکها،
قرقاولها و
مرغهای مگس
خوار، تنوع
خاصی از
رنگها،
الگوها، آرایشها
و زینتها را
به ماده ها
نشان می دهند،
در صورتیکه
ماده های گونه
های مختلف به یکدیگر
بسیار شبیه
هستند (Futuyma, 1998).
بنابر
این جدایی تولید
مثلی مهم است
زیرا از ادغام
گونه های
مختلف با یکدیگر
جلوگیری می
کند.


REFERENCE
Futuyma,
D. J., 1998. Evolutionary Biology. Sinauer
Associates, Inc.
کلادیستیک
سیستماتیک
فیلوژنتیک یا
تاکسونومی فیلوژنتیک
نیز نامیده می
شود و روشی
فرضی از روابط
بین ارگانیسم
ها است به
عبارت دیگر
روشی از
بازسازی درخت
های تکاملی
است که به وسیله
Will Hening(حشره
شناس)در سال 1950
توسعه یافت و
در 1980 پذیرفته
شد.این سیستم
طبقه بندی در
مقابل سیستم
طبقه بندی لینه
ای قرار می گیرد
پایه ی آنالیز
کلادیستیک
اطلاعاتی از ویژگی
ها یا صفات
موجودات است
که مورد توجه
ما است که این
ویژگی ها می
تواند فیزیولژیکی،آناتومیکی،رفتاری
یا سکانس های
ژنتیکی باشد
در نهایت نتیجه
آنالیزها یک
درخت است که
روابط میان
ارگانیسم ها
را نشان می
دهد مهم ترین
ویژگی کلادیستیک
عینی بودن آن
است (یعنی سلیقه
افراد در نتیجه
آنالیز موثر نیست
و با داده های یکسان،
همیشه یک پاسخ
به دست خواهد
آمد).
نام
کلادیستیک از
واژه کلاد(در یونانی
به معنای
شاخه)گرفته
شده است؛در
فارسی می توان
به جای این
واژه ،از واژه
یونانی تبار
استفاده کرد.یک
تبار گروهی از
جانداران است
که همگی از یک
نیای مشترک
اشتقاق یافته
اند،اگر و
تنها اگر تمام
جانداران
مشتق شده از
آن نیای
مشترک،زیر
مجموعه این
گروه قرار بگیرند.
در
مورد کلادیستیک
3 فرضیه وجود
دارد:
1- گروه
های موجودات
از یک جد
مشترک به وجود
آمده اند:این
فرضیه به وسیله
مدارک بسیاری
تایید می شود
و ضرورتا معنی
می دهد که همه
حیات بر روی
زمین مربوط به
یک جد مشترک
است.
2- در
دودمان یک
الگوی دو شاخه
شدن وجود
دارد:موارد بسیاری
این فرضیه را
مختل می کند
برای مثال بسیاری
زیست شناسان
نظریه ای که
چندین دودمان
از یک جمعیت
اصلی درهمان
زمان ناشی شده
اند را می پذیرند(مثل
ماهی های cichlid) یا
موضوعات دیگری
که بر ضد این
فرضیه است
قابلیت
زادآوری بین
گروههای مجزا
است که گاه بیگاه
در بعضی گروهها
اتفاق می افتد
(مثل گیاهان)
در واقع این
چنین
استثنائاتی
نادر هستند
بنابراین این
فرضیه صحیح
است.
3- تکامل
نتیجه پیشرفته
شدن صفات در
طول زمان
است:حالت اصلی
یا original
صفات را pelsiomorphic می
نامند که صفات
ابتدایی به
ارث رسیده از
جد مشترک
هستند و حالت
تغییر یافته
آن را apomorphic
می نامند که
مجموعه ی بی
نظیری از ویژگی
ها است که در
اجداد حضور
ندارد.
.یک
کلادوگرام یک
دیاگرام شاخه
ای ازگونه های
مشتق شده از
جد مشترک است
که توزیع و ویژگی
های مشتق شده
را نمایش می
دهند.کلادیستیک
اعتقاد دارد
که کلادوگرام
ها فرضیه های
قابل تستی از
روابط فیلوژنی
هستند.
همواره
فنتیک ها مورد
انتقاد کلادیستیک
ها واقع شده
اند زیرا نتایج
آنالیز
فنوگرام ها
لزوما مشابه
تاریخ تکاملی
موجودات نیست.در
مقابل فنتیک
ها ،آنالیز
کلادیستیک ها
بر روی صفات فیزیکی
وابسته نیست و
بر روی تاریخ
تکاملی تاکید
دارد در این
آنالیزها
ممکن است که
گروهها در ویژگی
های متعددی
مشترک باشند
برای مثال jelly fish sea star وhuman را
در نظر بگیرید.
jelly fishو
sea star هر
دو در اقیانوس
زندگی می کنند
تقارن شعاعی
دارند و بی
مهره هستند
بنابرآنالیزهای
فنتیکی در یک
گروه قرار می
گیرند و این
نتایج تکاملی
را بازتاب نمی
کند زیرا sea star از
لحاظ تکاملی
به انسان نزدیکتر
است تا jelly fish .بنابرین
کلادیستیک ها
روی صفات
مشترک تاکید
ندارند ولی بر
روی صفات مشتق
شده (آپومورفیک)
تاکید دارند
در مثال بالا
تقارن شعاعی،زیستگاه
آبی وساختار بی
مهره صفاتی
هستند که در
جد مشترک تمامی
حیوانات وجود
دارد و برای
تعیین روابط
آنالیز کلادیستیک
خیلی مفید نیستند
گروه
بندی کلادیستیک
باید
کاراکترهای زیر
را نمایش دهد:
1-همه ی گونه ها
در یک گروه
بندی باید از یک
جد مشترک مشتق
شوند
2-همه ی گونه های
مشتق شده از یک
جد مشترک در یک
تاکسون قرار می
گیرند.
اصطلاحات زیر
روش های
متفاوتی که در
گروه بندی میتواند
به کار رود را
شرح می دهد :
گروه
منوفیلتیك :
( Monopbyletic )
گروهی
كه شامل تمام
انواع مشتق
شده از یك
گونه اجدادی
باشد، به نوعی
كه هر گونهای
كه از نوع
اجدادی مذكور
تكامل یافته
باشد جزء آن
قرار گیرد و
تمام آن گونه
ها، از یك
گونهی اجدادی
و نه بیشتر
مشتق شده
باشند. مثلاً "پرندگان" گروهی
منوفیلتیك
است. در طبقه
بندی، تنها
گروههایی كه
منوفیلتیك
باشند از نظر
علمی
معتبراند
گروه پلی
فیلتیك ( Polyphyletic )
مجموعهای
از گونهها كه
از بیش از یك
گونهی اجدادی
حاصل شده
باشند به نوعی
كه جدیدترین
حد مشترك آنها
جزء این گروه
قرار نگیرد.
مثلاً اگر
پستانداران و
پرندگان را با
هم در یك گروه
ساختگی قرار
دهیم، این
گروه پلی فیلتیك
است. این چنین
گروهی غیر
قابل استناد و
مردود است.
گروه
پارافیلتیك (
Paraphyletic )
گروهی
كه شامل توالیای
از گونههای
رو به تكامل
به سمت یك
گروه نهایی
باشد، به طوری
كه یك یا چند
شاخهی نهایی
را ـ مثلاً به
دلیل تغییرات
شدید ظاهری ـ
كه از همان جد
مشترك مشتق
شده اند، جزء
آن محسوب
نكنند.. نمونه
بارز آن « ماهیها»
هستند. حتی
گروههای
شناخته شده تری
همچون «
خزندگان» نیز
پارافیلتیكاند
زیرا پرندگان
در حقیقات
ادامهی مسیر
تكامل
خزندگاناند.
مسلماً
گروههای
پارافیلتیك نیز
از نظر علمی
مردوداند و
باید در تعریف
آنها تجدید
نظر نمود.
مزیت های
دسته بندی فیلوژنتیکی:
این دسته
بندی 2 مزیت
اصلی بر سیستم
لینه ای
دارد:1-روابط
تکاملی
موجودات را
نشان می دهد.2-این
طبقه بندی
تلاش برای
رتبه بندی
موجودات
ندارد.در طبقه
بندی لینه ای
رتبه بندی
گروه های
ارگانیسم ها
به صورت
سلسله،شاخه،راسته
و....وجود دارد
که این موجب
گمراهی می شود
زیرا گروه های
متفاوت با این
رتبه بندی هم
ارزش می شوند .
رقابت می
تواند به
عنوان یک بر
هم کنش بین
ارگانیسم ها یا
گونه ها در این
جهت که یک
گونه یا ارگانیسم
شایستگی بیشتری
برای حضور در
طبیعت داشته
باشد، تعریف
شود.
ریشه لاتین
فعل compete ، competere است که به
معنای نزاع
کردن با یکدیگر
می باشد.
رقابت به طور
طبیعی بین
ارگانیسم های
زنده که به
طور همزمان در
محیط های یکسان
وجود دارند رخ
می دهد. برای
مثال جانوران
برای بدست
آوردن بیشتر
آب، غذا، جفت
و غیره با یکدیگر
رقابت می
کنند. رقابت
ممکن است بین 2
عضو از یک
گونه تحت
عنوان رقابت
درون گونه ای ( intraspecific )
رخ دهد در حالی
که سایر رقابت
ها می تواند
گونه های
مختلف را درگیر
کند که تحت
عنوان رقابت بین
گونه ای ( interspecific )
بدان اشاره می
گردد.
رقابت درون و
بین گونه ای
از عنوان های
مهم در اکولوژی
می باشند. به
طور کلی رقابت
یکی از
فاکتورهای biotic و abiotic
مؤثر بر
ساختار
اجتماع است.
رقابت بین
اعضای یک گونه
( درون گونه ای )
نیروی محرک
پشتیبان
تکامل و
انتخاب طبیعی
است. رقابت
برای منابعی
همچون غذا،
آب، قلمرو و
نور خورشید رخ
می دهد که
سرانجام تسلط
و بقای بهترین
گونه های
موجود را نتیجه
می دهد. در
رقابت بین
گونه ای میزان
محدودی از
منابع در
دسترس است و
چندین گونه
ممکن است به این
منابع وابسته
باشند. هر
کدام از این
گونه ها برای
کسب سهم بیشتری
از این منابع
با هم رقابت می
کنند.
بر طبق تئوری
تکاملی رقابت
درون و بین گونه
ای، نقشی
بحرانی را در
انتخاب طبیعی
ایفا می کند.
برای مثال یک
درخت کوتاه
نور خورشید
کمتری نسبت به
درخت مجاور
بلندتر دریافت
می کند.
انواع
رقابت
این واژه ها
مکانیسم هایی
که رقابت از
طریق آن ها رخ
می دهد را شرح
می دهند که
عموماً می
تواند به 2 مسیر
مستقیم و غیر
مستقیم تقسیم
گردد. این
مکانیسم ها
برای هر 2 نوع
رقابت بین
گونه ای و
درون گونه ای
به کار می
روند :
رقابت
تداخلی ( interference
)
رقابت تداخلی
شامل بر هم
کنش های مستقیم
بین افراد
همانند جنگ بر
سر منابع
محدود است. این
شکل از رقابت (
گرچه نه همیشه
) برای هر 2 فرد یا
گونه درگیر
مضر است. برای
مثال شکارچیانی
همانند canids
مزیت بزرگی
اندازه نسبت
به شکارچیان
کوچک تر
همانند foxs
و weasels را
دارند. یک بر
هم کنش سخت و
شدید میان 2
فرد از این گونه
ها پیروزی
شکارچی بزرگ
تر را در پی
خواهد داشت.

شکل. رقابت بین
2 گوزن قرمز نر
مثالی از
رقابت تداخلی
درون گونه ها
می باشد.

شکل. رقابت
تداخلی بین
گونه ای بین شیرها
و کفتارها
رقابت
استثماری ( exploitation
)
رقابت
استثماری فرمی
از رقابت است
که طی آن یک
گونه به طور
مؤثر استفاده
بیشتری از یک
منبع داشته و
باعث کاهش آن
می گردد به
طوری که دسترسی
منبع برای سایر
گونه ها را از
بین می برد. یک
مثال مناسب از
این نوع رقابت
در گونه های aphid یافت شده
که برای شیره
فلوئم گیاه
رقابت می
کنند. هر گونه aphid که از شیره
گیاه میزبان
تغذیه می کند،
پس مانده کمی
برای گونه های
رقیب باقی می
گذارد.
رقابت
آشکارا ( ظاهری
)
این نوع
رقابت مستقیماً
بین 2 گونه رخ می
دهد که هر 2 به
وسیله شکارچی یکسان
صید می شوند.
برای مثال
گونه های A
و گونه های B هر 2 برای
شکارچی C
طعمه محسوب می
گردند. افزایش
گونه های A
باعث کاهش
گونه های B می شود زیرا
افزایش گونه
های A
تعداد گونه های
C
شکارچی را
افزایش خواهد
داد بنابراین
تعداد بیشتری
از افراد گونه
های B
شکار خواهند
شد.
رقابت
درون گونه ای
رقابت درون
گونه ای شکل
خاصی از رقابت
است که در آن
اعضای گونه های
مشابه برای
منابع طبیعی
مشابه ( مثل
غذا، نور،
مواد مغذی و
فضا ) در یک
اکوسیستم با
هم رقابت می
کنند. این شکل
از رقابت
برخلاف رقابت
بین گونه ای می
باشد که در آن
گونه های
مختلف با هم
رقابت می
کنند.
برای مثال 2
درخت از یک
گونه مشابه که
در کنار یکدیگر
رشد می کنند
برای نور، آب
و مواد مغذی
موجود در خاک
با هم رقابت
خواهند داشت.
با دریافت
منابع کمتر آن
ها بسیار ضعیف
خواهند شد. (
برای مثال رشد
کمتر و میزان
تولید دانه
کمتر ) بنابراین
درختان خود را
با رشد طویل
تر ( بلند تر
شدن ) یا تکامل
سیستم ریشه ای
بزرگ تر از طریق
انتخاب طبیعی
سازش می دهند.
ملخ مثالی دیگر
در این مورد می
باشد. با
خوردن علف هر
ملخ خود را از
هر نوع غذای دیگری
بی بهره می
سازد. این
نمونه ای از
رقابت
استثماری ( explotation ) است بدان
معنا که ملخ
ها به طور
مستقیم بر یکدیگر
اثر نمی کنند
بلکه به وسیله
اثر بر منابع
آن ها ( در این
مورد علف ) اثر
منفی بر روی
رشد سایرین و
تولید مثل آن
ها دارند. در
مثال دیگر
رقابت درون
گونه ای ممکن
است نمونه ای
از رقابت
تداخلی باشد
که در آن
جانوران مستقیماً
بر روی یکدیگر
اثر می
گذارند.
این نمونه ای
از ویژگی
برجسته
جانوران خشکی
زی است. یک فرد
به طور فعال
با استثمار
کردن منابع که
معمولاً غذا و
فضا است مانع
فعالیت سایرین
می گردد.
رقابت
بین گونه ای
رقابت بین
گونه ای در
اکولوژی شکلی
از رقابت است
که طی آن افراد
گونه های
مختلف برای
منابع یکسان
در یک اکوسیستم
( از جمله فضای
زندگی یا غذا )
رقابت می
کنند. اگر یک
درخت در یک
جنگل انبوه و
متراکم
بلندتر از
درختان اطراف
رشد کند این
درخت قادر به
جذب بیشتر نور
خواهد بود و
نور کمتری در
دسترس درختان
موجود در زیر
سایه درخت بلندتر
قرار خواهد
گرفت. این نوعی
رقابت بین
گونه ای است. یک
مثال در بین
جانوران می
تواند مورد چیتا
ها و شیرها
باشد. از آن جایی
که هر 2 گونه از
طعمه های
مشابه تغذیه می
کنند آن ها به
طور منفی به
وسیله حضور یکدیگر
تحت فشار قرار
می گیرند زیرا
آن ها غذای
کمتری خواهند
داشت. برخی
مواقع شیرها
طعمه کشته شده
به وسیله چیتا
ها را می
دزدند. رقابت بین
گونه ای ممکن
است زمانی رخ
دهد که افراد 2
گونه جدا یک
منبع محدود در
نواحی یکسان
را به اشتراک
دارند.

شکل.
درختان در این
جنگل بنگلادشی
برای ذریافت
نور رقابت بین
گونه ای
دارند.
(هم تکاملی)
تغییردرترکیب
ژنثیکی یک
گونه (یا گروه)
،در جواب به یک
تغییر ژنتیکی
در گونه دیگری
میباشد.در یک
نگاه اجمالی
ممکن است اینطور
تصور شود که، هم تکاملی، همه چیز
را در بر می گیرد.این
فرض ممکن است
از این حقیقت
که همه
موجودات زنده
باهم در
تعاملند واحتمالأ
تکاملشان به
طرقی برهم تاثیر
میگذارد نشأت
گرفته باشد.
اصطلاح هم تکاملی، برای
توصیف مواردی
که دویا چند
گونه بطور
متقابل تکامل
همدیگر را تحت
تاثیر قرار میدهند
استفاده میشود
برای مثال یک
تغییر تکاملی
در مورفولوژی یک
گیاه ممکن است
مورفولوژی گیاهخواری
که از آن گیاه
تغذیه میکند
را تحت تاثیر
قرار دهد که
در نتیجه
تکامل ایندو
تحت تاثیرهمدیگر
قرار میگیرد.
احتمالأ
زمانیکه گونه
های مختلف
برهمکنشهای
اکولوژیکی نزدیکی
با یکدیگر
دارند، هم تکاملی رخ می
دهد. این
روابط اکولوژیکی
عبارتند از :
1.
شکارچی/شکار
و انگل/میزبان
2.
(Competitive ) 2.گونه
های رقابتی
3.
(Mutualistic)3.گونه
های همزیست
هم
تکاملی روشهای
گوناگونی
دارد.در برخی
از
موارد،کاملأ
اختصاصی
است.مثلأ بین
دو فعالیت
سلولی؛
کلرپلاست ومیتوکندری
زمانی بصورت
تک سلولی
وآزاد زندگی می
کردند،بعدأ
وارد سیتوپلاسم
سلولهای دیگر
شدند.(مثالی
از ) endosymbiosis پدیده
روش
دیگر شامل
برهمکنش
رقابتی بین دو
گونه ویژه می
باشد.یک مثال
خوب می باشد؛Plethodon مطالعه
روی سمندر
دوگونه
از این سمندر
شدیدأ باهم
رقابت می
کنند. Great Smoky در
کوههای
(اگر میزان
جمعیت هر گونه
کاهش یابد،گونه
دیگر جمعیتش
افزایش می یابد)
در اینجا هم یک
برهمکنش
دوجانبه بین
دوگونه وجود
دارد.
گیاهان
وحشرات نمونه
های کلاسیکی
از هم تکاملی
را نشان می
دهند.بسیاری
از گیاهان و
گرده افشانهایشان
کاملأ به هم
وابسته
ومنطبقند
وروابطشان بسیار
منحصر به فرد
است وزیست
شناسان این
جوربودن را در
نتیجه فرایند
هم تکاملی میدانند.(coevolution)

:
(تکامل
همگرا)
تکامل
سازگار
ساختارهای به
ظاهر مشابه
مثل بال
پرندگان
وحشرات،در گونه های نامرتبط
،که در معرض
محیط های یکسان
قرار
دارند.(تکامل
همگرا).
دانشمندان
علم سیستماتیک
در تلاشند تا
بوسیله بررسی
شباهتهای بین
موجودات زنده،ارتباطات
تکاملی ایجاد
کنند.اما
متأسفانه،
همه شباهتها
از یک جد
مشترک به ارث
نمی رسند.اگر
گونه ها دارای
نقشهای
اکولوژیکی
وانتخاب طبیعی
یکسان باشند،
که به شکل
سازگاریهای
مشابه وقابل
مقایسه
درآمده
باشد،هرچند
از شاخه های
تکاملی مختلفی
باشند، ولی
ممکن است شبیه
به هم به نظر
برسند.این را
تکامل همگرا می
نامند و تشابه
در نتیجه
همگرایی را
آنالوگی گویند.
در
علوم طبیعی
؛تکامل
همگرا،به
موجودات زنده
ای که رابطه
نزدیکی باهم
ندارند،مربوط
می شود که هر
کدام مستقلأ
صفات مشابهی
کسب می کنند
درحالیکه در
اکوسیستم های
جدا یا گاهأ مختلفی
قرار می گیرند.

مثال:
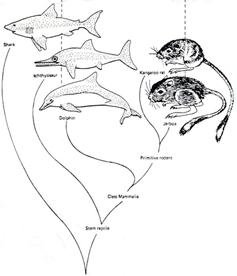

: تکامل
همگرا در مهره
داران
تحت
تنشها
وفشارهای محیطی
مشابه،ممکن
است موجودات
زنده ایکه به
ظاهر شبیه یکدیگرند،ولی
از اجداد
مختلفی میباشند،
پدیدار شوند.
همگرایی
در فرم بدن
مهره داران :

در سراسر
بدن،در تکامل
مهره داران
مشترک است.
مثال زیر؛(در
سمت چپ) یک
پستاندار،یک
خزنده ویک ماهی
را که همه
شکارچیان اقیانوسی
شده اند، و(در
سمت راست) دو
جوند صحرایی
شکارچی
حشره،که با شیوه
زندگی جهش سریع،
سازگار شده
اند را نشان می
دهد.
3- Parallel
* تکامل صفات
مشابه در
گروههای
مختلف موجودات
زنده.
* شبیه
بودن به یا
آنالوگ بودن
با:ادعا شده
است که تکامل
جنینی موازی
تکامل گونه ها
می باشد.
(تکامل
همسو)Parallel
evolution*
تکامل
مستقل صفات
مشابه می باشد
که از یک نقطه
اجدادی
مشابه، در نتیجه
محیط های یکسان
یا فشارهای دیگرتکاملی
شروع می
شود.غالبأ این
حالت در اجدادی
که دارای
ارتباط نزدیکی
هستند می
باشد، جایی که
چندین گونه به
طرق یکسان، به
تنشهای محیطی،
پاسخ مشابه می
دهند.
* تکامل
گروههایی که
بطور جغرافیایی
جدانشده اند
ودارای
شباهتهای فیزیکی
هستند.یک مثال
برجسته،شباهت
بین
پستانداران
مارسوپیال
استرالیا
وپستانداران
جفت دار،درجایی
دیگر می باشد
که هرچند مسیرهای
تکاملی
جداگانه ای دارند ولی
فرمهای مشابه
قابل توجهی
دارند
(
تکامل واگرا)
* فرایندی
که طی آن یک
گونه به دو یا
چند زاده یا
اشکال متفاوت
درمی آید،
همچنین روند
ردیابی(دنبال
کردن) دو یا چند
گونه به
عقب،برای رسیدن
به یک جد
مشترک را گویند.
* انبوهی
از اختلافات بین
گروهها،که می
تواند منجر به
تشکیل یک گونه
جدید شود،که
معمولأ در نتیجه
انتشار گونه
های یکسانی که
با محیط های
متفاوتی
سازگار شده
اند، می باشد. یعنی
در این گونه
جدید جهش های
ویژه مفیدی ایجاد
شده که منجر
به انتخاب طبیعی
آنها گردیده
است.
* نوعی
تغییر تکاملی
ا ست که منجر
به افزایش
اختلافات
مورفولوژیکی
بین اجداد اولیه
که به هم شبیه
بودند، می
شود.
روندی که طی
آن دو گونه
برای سازگاری
بیشتر با محیطشان
از صفاتی که
از یک جد
مشترک گرفته
اند، دور می
شوند.

به
عنوان مثال
اگر تنوع بین
گونه های
پرندگان را
تصور کنیم؛اردکها
،مرغ مگس
خوار،شتر مرغ
و پنگوئن،همه
از یک گونه پرنده
اجدادی نشأت
گرفته اند و
تا این
زمان،همه
آنها به خاطر
سازگار شدن با
محیط های خاص
خود، ویژگیهای
خاصی یافته
اند که در نهایت
منجر به دور
شدن آنها از
همدیگر شده
است.
تکامل
واگرا می
تواند برای
تشخیص صفات،
در بیولوژی
ملکولی بکار
برده
شود،مثلأ از
تکامل واگرا می
توان برای
ژنها و پروتئینها
استفاده
کرد،مثل توالی
های نوکلئوتیدی
و پروتئینی که
از دو یا چند
ژن همولوگ
مشتق شده
اند.هم ژنهای
ارتولوگ(که نتیجه
روند گونه زایی
می باشند) و هم
ژنهای
پارالوگ (که
نتیجه
همانندسازی
ژنی درون یک
جمعیت می
باشند) تکامل
واگرا را نشان
می دهند. ممکن
است تکامل
واگرا به علت
دومی (ژنهای
پارالوگ)، بین
دو ژن در یک
گونه رخ دهد.
Exaptation در
لغت به این
مفهوم می
باشد:
کاربرد یک
ساختار یا عمل
بیولوژیکی به
منظوری
متفاوت از آن
چیزی که ابتدا
به وجود آمده
است، به عبارت
دیگر عملکرد یک
صفت می تواند
تحت تأثیر
انتخاب طبیعی
به گونه ای تغییر
پیدا کند که
صفتی که ابتدا
کاری را انجام
می داده است
بعد از گذشت
مراحلی به عمل
دیگری اختصاص
می یابد. این
پروسه که هم
در آناتومی و
هم در رفتار
مشاهد می شود
توسط Gould و , Urba Exaptation نامیده
شده است.
تاریخ:
انتخاب
طبیعی نمی
تواند آینده
را پیش بینی
کند اما
ساختمانهای
موجود برای
استفاده های
جاری را بهبود
می بخشد برای
مثال:
استخوانهای
مشبک و پرهای
پرندگان به
عنوان سازگاری
برای پرواز
تکامل نیافته
اند.
-
در
ابتدا آنها باید
برای اجداد
پرندگان که
خزنده دوپا
بودند مفید
بوده باشند و
در طول تغییرات
برای پرواز به
کار برده
شدند.
-
Exaptation نمی
تواند اثبات
شود ولی برای
اینکه چگونه
طرح های جدید
به تدریج می
توانند در طی یک
سری مراحل
واسطه ای به
وجود آیند،
توضیحی ارائه
می دهد که هر
کدام از این
طرح ها کاربردهایی
در ارگانیسم
دارد.
-
تکامل و
طرح های
نوظهور توسط
بازسازی
ساختارهای قدیمی
برای اعمال جدید،
عقیده داروین
را منعکس می
سازد که در آن
انباشتگی تغییرات
کوچک در اثر
انتخاب طبیعی
باعث بوجود
آمدن تغییرات
بزرگ می شود.
-
این
ایده که کارآیی
و عملکرد یک
صفت ممکن است
طی تاریخ
تکاملی اش
توسط Charles Darwin
شروع شد. چندین
سال این پدیده
به عنوان Preadaptation نامگذاری
شده بود.
متأسفانه این
واژه به اندیشه
قبلی اشاره می
کند که با
اصول اولیه
انتخاب طبیعی
مغایر است. این
ایده توسط
محققین معتددی
بررسی شده
بود. در سال 1982 Gouldو Urbaواژه
Exaptation را
مطرح کردند.
متأسفانه بحث
بعدی این تعریف،
دو مقوله با
مفاهیم
متفاوت برای
نقش "سازگاری"
داشت:
1- صفتی که
در گذشته توسط
انتخاب طبیعی
برای یک عمل ویژه
شکل گرفته بود
(Aadaptation) برای یک
کاربرد جدید
در نظر گرفته
می شود – Cooptation(مثل
اکولوکیشن در
خفاش)
2- صفتی که
علت آن به
عملکرد مستقیم
انتخاب طبیعی
نسبت داده نمی
شود برای
کاربرد جدید
در نظر گرفته
می شود.
پرها
مثال کلاسیک Exaptation هستند
پرها در اصل
فلس بودند که
بعد از مراحلی
به سیستم
کنترل دمایی
تغییر می یابند.
توانایی حفظ
دما در یک روز
گرم سیستم خنک
کننده بسیار
مناسبی ایجاد
می کند،
بنابراین
فشار انتخاب
طبیعی روی
پرهای حیوانات
اینچنین است
که قادر به
حرکت هوا به
طور کاربردی و
موثرتری شوند.
در نتیجه
اندام حرکتی
جلویی این حیوانات
که اساسا برای
جابجایی به
کار برده می
شد تحت Exaptation به
عنوان سیستم
خنک کننده به
کار می رود
سرانجام طی
مراحلی این سیستم
های خنک کننده
بسیار موثر و
کارا
توانستند به
مدت چند ثانیه
برای پرواز
کردن به کار
برده شوند و
به این ترتیب
پرواز کننده
های خوب
توانستند از
دسترس شکارچی
فرار کرده و
سرانجام ژنهایشان
را به نسل بعدی
منتقل کنند. پس
در عرض صد میلیون
سال پوشش به
عنوان کنترل
دمايي و سپس سیستم
کنترل دمایی
به عنوان سیستم
پروازی تغییر یافته
است
دستان
انسان نیز از یک
توالی طولانی Exaptation حاصل
می شود و نیز
باله ها در
ماهی؛ اندام
حرکت جلویی در
فرم های
انتقالی
مانند ,Tiktaalik Acanthostega یا Ichthyostegaاندام جلویی
در تتراپودا؛
ساختارهایی
برای گرفتن
شاخه های درخت
در پریماتهای
درختی
ساختارهایی
برای چنگ زدن
در همونیدها و
انسانها
Founder Effects and Bottlenecks
هر جمعیتی
ممکن است محدودیت
هایی در
اندازه داشته
باشد که به آن bottleneck یا
تنگنا گفته می
شود. یکی از این
تنگناهای
جالب زمانی ایجاد
می شود که
قرار باشد جمعیت
جدیدی توسط
چند موسس که
گاهی یک جفت
تولید مثلی
هستند ،ایجاد
شود.

مثال:
(فوک
های شمالی ) Northern elephant seals
به دلیل
کشتار توسط
انسان در دهه 1890
این اثر را
نشان می دهد .

رانش ژنتیکی
تصادفی که در
این جمعیت ایجاد
می شود را
اصطلاحا اثر موسس
founder effect مینامیم
. در یک چنین
جمعیتی
معمولا آلل های
فراوان وجود
ندارد. با این
حال هتروزیگوتی
در نسل اول
هنوز خیلی
کاهش پیدا
نکرده است . در
کلنی که توسط یک
جفت موسس ایجاد
شده باشدN=2
بوده و طبق
فرمول)H ![]() که
H هتروزیگوتی
در جمعیت مبدا
است ،پس هتروزیگوتی
در نسل اول 25
درصد کاهش پیدا
میکند.چنانچه
تعداد افراد
کلنی سریعا
افزایش پیدا
کند ، از بین
رفتن تنوع ژنتیکی
آهسته خواهد
بود ، اما اگر
جمعیت کوچک
باقی بماند
رانش ژن موجب
کاهش تنوع ژنتیکی
خواد شد .
که
H هتروزیگوتی
در جمعیت مبدا
است ،پس هتروزیگوتی
در نسل اول 25
درصد کاهش پیدا
میکند.چنانچه
تعداد افراد
کلنی سریعا
افزایش پیدا
کند ، از بین
رفتن تنوع ژنتیکی
آهسته خواهد
بود ، اما اگر
جمعیت کوچک
باقی بماند
رانش ژن موجب
کاهش تنوع ژنتیکی
خواد شد .
مثال:
جمعیت اصلی در سمت چپ و جمعیت موسس در سمت راست است .

Founder effect
وقتي رخ
مي دهد كه يك
جمعيت جديدي
به وسيله ي تعداد
كمي از افراد
از جمعيت اصلي
ايجاد شودكه در
اين جمعيت
كوچك تغيير
پذيري ژنتيكي
كاهش مي يابد. بنابر اين
Founder effect
يك حالت خاص
از دريفت
ژنتيكي است كه
جمعيت جديد با
تعداد افراد
بسيار كمتربه
طور مشخصي هم از
نظر ژنتيكي و
هم از نظر
فيلوژنتيكي
از جمعيت
والدي كه از
آن مشتق شده
است متمايز مي
شود. اين جمعيت
جديد يك
افزايش درون
زادآوري و كاهش
تغيير پذيري
ژنتيكي را
نشان مي دهد.
در
حالت حداكثر Founder effect منجر
به گونه زايي
و تكامل
متعاقب گونه
هاي جديد شود.

مثال:
جمعيت Amish در ايالات
متحده امريكا
كه يك جامعه
كوچكي را تشكيل
مي دهندكه
سندروم Ellis-van
Creveld در
بين اين
اجتماع بسيار
معمول است(يك
انگشت اضافي
در دست و
نارسايي قلبي
دارندوكوتاه
قد هستند.)كه
يك Founder
effect
مي باشد
Population bottlenecks( genetic bottlenecks)
يك
رويداد
تكاملي است كه
در آن درصد
قابل توجهي از
يك جمعيت كشته
ميشوند(به
وسيله ي شكار
شدن ويا
بلاياي طبيعي
مثل سيل و
خشكي) ويا به
طريقي از
توليد مثل
آنها جلو گيري
مي شود. از
آنجايي كه
ميزان دريفت
ژنتيكي با
اندازه جمعيت
نسبت معكوس
دارد بنابر
اين جمعيتbottleneck دريفت
ژنتيكي را
افزايش ميدهد
ودر آنها به
دليل كاهش
منابع جفت
گيري درون زاد
آوري(( inbreedingافزايش مي
يابد.

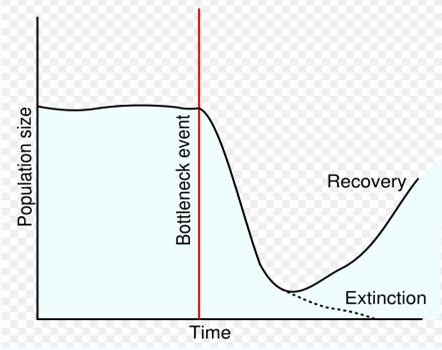
مثال1:
فيل هاي
آبي شمالي كه
جمعيت انها به
حدود 30 در دهه 1860رسيد
اگرچه اكنون
تعداد آنها
هزاران
ميليون است كه
علت كاهش
جمعيت آنها در
دهه 1860به دليل
باتل نكي بود
كه انسان ها
به وسيله ي
شكار آنها به
آنان تحميل
كردند.اين
جمعيت از فيل
هاي آبي دگرگوني
ژنتيكي كمتري
از فيل هاي
آبي جنوبي كه
شكار نشدند
نشان مي دهند
فيل هاي
آبي نجات
يافته از شكار
به مرور آفزايش
يافتند ويك
جمعيت جديد را
به
وجودآوردند.

مثال2
در مورد
جمعيت گراز
آمريكايي كه
فقط جمعيت كمي
از گراز ها با
ژنوتيپbbاز
حوادث طبيعي
نجات يافتند
كه اينها پايه
گذار يك جمعيت
جديد از گراز
ها شدند كه
گنجينه ژني
آنها خيلي از
جمعيت اصلي
متفاوت مي
باشد.

جدایی
گامتی
چنانچه
میدانیم
مفهوم بیولوژیک
گونه بر پایه ی
جدایی تولید
مثلی استوار
است وجدایی
تولید مثلی شامل
تمام
فاکتورهایی
است که از تولید
هیبرید های زایا
و زیستا توسط
دو گونه ی
مختلف جلوگیری میکند.
سدهای پیش زیگوتی
و پس زیگوتی
خزانه ی ژنی
گونه های بیولوژیک
را ازهم جدا میکند.از
سدهای پیش زیگوتی
میتوان به جدایی
زیستگاهی،رفتاری،مکانیکی
و گامتی اشاره
کرد که از این
میان تنها جدایی
گامتی است که
در شرایط آزمایشگاه
و با انتخاب
مصنوعی قابل
رفع نیست.جدایی
گامتی سبب
ناسازگاری میان
اسپرم و تخم
گونه های
مختلف میشود.در
این مکانیسم
گامت ها به دلیل
آنکه قادر به
تشخیص یکدیگر
نیستند با هم
ادغام نمی
شوند.شناسایی گامت
ها ممکن است
بر پایه ی
وجود مولکول
های ویژه روی
پوشش اطراف
تخم باشد که
تنها به مولکول
مکمل خود روی
اسپرم فرد هم
گونه اش می
چسبد.چنین
مکانیسم
شناسایی مو
لکولی است که یک
گل را قادر می
سازد تا گرده ی
همان گونه را
از گرده ی
گونه ی دیگر
تشخیص دهد .
جدایی
گامتی به
فاکتورهای زیادی
وابسته است که
از آن جمله می
توان به موارد
زیر اشاره
کرد:
1.
ممکن است
اسپرم فاقد
آنزیم های
مناسب و کافی
برای عبور از
لایه های تخم
باشد
2.اسپرم
فاقد پروتئین
مکمل با
مولکول های
سطحی تخم باشد
3.تعداد
کروموزوم ها
در گامت های
دو گونه با هم
برابر نباشد. این
مسئله مخصوصا
در حیوانات که
وجود دو گامت
هاپلوئید با
تعداد
کروموزوم
مساوی جهت
لقاح الزامی
است نمود پیدا
می کند.جدایی
گامتی به خصوص
در محیط های
آبی مطرح می
شود.زیرا بسیاری
از حیوانات
آبزی گامت های
خود را در آب
رها می کند تا
لقاح انجام گیرد.برای
مثال میتوان
به حلزون دو
کفه ای اشاره
کرد در این
حلزون
همانطور که در
تصویر می بینید
جنس ماده توده
ای عظیم از
تخم را از سیفونش
رها می
کند.جنس نر نیز
میلیون ها
اسپرم در آب
آزاد می کند.تخم
ها و اسپرم ها
در آب رها میشوند
تا همدیگر را
پیدا کنند.در
این زمان اگر
حلزونی از
گونه ای دیگر
در حال آزاد
سازی گامت های
خود باشد این
گامت ها نیاز
به مکانیسم
مطمئنی برای
شناسلیی گامت
فرد هم گونه ی
خود دارند و
لذا از پروتئین
های سطحی ای
که بطور دقیق
با مو لکول
مکمل خود
ادغام می شوند
استفاده می
کنند.

Here
you see a giant clam releasing a huge cloud of eggs from its siphon.
در
موجوداتی نظیر
انسان که تولید
مثل داخلی
دارند دستگاه
تناسلی ماده
حتی نسبت به
اسپرم فرد هم
گونه اش بسیار
غریبه ناپذیر
است.و اسپرم
پس از ورود به
واژن با سدهای
بسیاری من
جمله ترشحات
موکوزی رو به
رو میشود
شواهدی وجود
دارد که فرد
ماده علیه
اسپرم آنتی
بادی تولیدمیکند
حتی اگرا سپرم
خود را به
تخمک برساند
هنوزبه شانس زیادی
برای انجام
لقاح نیاز
دارد زیرا مو
لکول های
شناسا گر گامت
ماده می تواند
تنها به
مولکول مکمل
اسپرم از همان
گونه بچسبد.جدایی
گامتی میتواند
در میان دو
جمعیت که نزدیک
هم زندگی میکنند
اتفاق بیفتد.
این فرایند میتواند
به عنوان یک
مکانیسم جدایی
مطرح گردد که
در مراحل اپتدایی
گونه زایی
اتفاق می
افتد.
رانش
ژنتیکی:
رانش
ژنی یا رانش
اللی تغییر در
فراوانی نسبی
اشکال مختلف یک
ژن(الل های ژن)
است که در یک
جمعیت اتفاق می
افتد و از این
واقعیت منشا می
گیرد که الل
ها در
فرزندان، یک
نمونه تصادفی
از الل ها در
والدین هستند
ونیز به دلیل
نقش شانس و
تصادف در تعیین
این که ایا یک
فرد معین باقی
می ماند و تولید
مثل می کند
.رانش ژنی
ممکن است سبب
از بین رفتن
گوناگونی های
ژنی به صورت
کامل شود و
بنابراین
تنوع ژنتیکی
را کاهش می
دهد.
رانش
ژنی یکی از
پروسه های
تکاملی تدریجی
است که تغییرات
در فراوانی
الل ها را هدایت
می کند.بر
خلاف انتخاب
طبیعی تغییراتی
که معلول رانش
ژنی هستند به
وسیله فشارهای
سازشی و محیطی
به وجود نیامده
اند وممکن است
برای موفقیت
تولیدمثلی مفید،
مضر یا بی اثر
باشند
تاثیر
رانش ژنی در
جمعیت های
کوچک بیشتر و
در جمعیت های
بزرگ کمتر
است.در بین
دانشمندان
بحث های شدیدی
در مورد اهمیت
نسبی رانش ژنی
در مقایسه با
انتخاب طبیعی
وجود داشته
است.رونالد فیشر
دیدگاهی را به
وجود اورد که
در ان رانش ژنی
نقش کوچکی را
در تکامل بازی
می کرد و این دیدگاه
برای چندین
دهه رایج بود.
در سال 1968, Matoo Kimura
باتوجه به
تئوری اش
"تئوری خنثی
تکامل مولکولی"
بحثی را به
راه انداخت که
ادعا می کرد بیشترین
تغییرات در
ماده ژنی از
رانش ژنی نتیجه
شده است.
رانش
ژنی علیه
انتخاب طبیعی:
اگرچه
هر دو پروسه
تکامل را پیش
می برند رانش
ژنی به صورت
تصادفی عمل می
کند در حالی
که انتخاب طبیعی
غیر تصادفی
است.به این
خاطر که
انتخاب طبیعی
نشان دهنده بر
همکنش اکولوژیکی
یک جمعیت است
و رانش به
عنوان یک روش
نمونه برداری
در نسل های
متمادی بدون
توجه به
فشارهای شایستگی
تحمیلی به وسیله
محیط در نظر
گرفته می
شود.در حالی
که انتخاب طبیعی
جهت دار است و
تکامل را به
وسیله سازش های
محیطی قابل
توارث پیش می
برد رانش ژنی
جهت ندارد و
تنها توسط
شانس و احتمال
ریاضی هدایت می
شود.
به
عنوان یک نتیجه،
رانش بر روی
فراوانی های
ژنوتیپی در
داخل یک جمعیت
بی توجه به
ارتباطشان با
فنوتیپ عمل می
کند.تغییرات
ژنوتیپی توسط
رانش ممکن است
منجر به تغییرات
فنوتیپی شود ویا
ممکن است تغییر
فنوتیپی به
وجود نیاورد.در
رانش هر الل
در یک جمعیت
به صورت تصادفی
و مستقل تحت
تاثیر واقع
شده است.رانش
در قبال الل
های مفید یا
مضر،به صورت
کور و نابینا
عمل می
کند.سرانجام
انتخاب طبیعی
به طور مستقیم
روی فنوتیپ و
به صورت غیرمستقیم
روی ژنوتیپ
عمل می
کند.انتخاب به
طور ویژه به
سود یا زیان
سازشی ارئه
شده توسط یک
صفت فنوتیپی
واکنش می دهد
و بنابراین به
شیوه متفاوتی
بر روی ژن ها تاثیر
می
گذارد.انتخاب
به طور غیرمستقیم
به الل هایی
که فنوتیپ های
سازشی مفید را
به وجود می
اورند پاداش می
دهد.به این
صورت که افزایش
موفقیت تولیدمثلی
این فنوتیپ ها
باعث افزایش
فراوانی الل
های ان ها می
شود.
در
جمعیت های طبیعی
رانش ژنی و
انتخاب طبیعی
به تنهایی عمل
نمی کنند همیشه
هر دو نیرو
نقش
دارند،اگرچه
الل ها به میزان
متفاوتی از
رانش ژنی و
انتخاب طبیعی
متاثر می شوند
و این به
اندازه جمعیت
بستگی
دارد.زمانی که
جمعیت ها خیلی
کوچک هستند
رانش غالب
خواهد بود و
ممکن است الل
های مطلوب را
حذف والل های
غیر مطلوب را
حفظ کند.
اثرات انتخابی
ممکن است دیده
نشوند، چون تغییرات
کوچکی که در
فراوانی الل
ها به وجود می
اورند، توسط
رانش تحت
الشعاع قرار می
گیرد.
در
جمعیت های
بزرگ اشتباه
نمونه برداری
کوچک است و حتی
در پی نسل های
متعدد تغییرات
کمی در فراوانی
الل ها مورد
انتظار است.حتی
فشارهای
انتخابی ضعیف
که روی الل ها
عمل می
کنند،فراوانی
ان ها را به
طرف بالا یا
پایین تغییر می
دهند.(بسته به
این که الل ها
مفید یا مضر
باشند.)
البته
در مواردی که
فراوانی الل خیلی
کوچک است رانش
می تواند حتی
در جمعیت های
بزرگ بر
انتخاب طبیعی
پیروز شود.برای
مثال در حالی
که موتاسیون
های غیر مفید
در جمعیت های
بزرگ به سرعت
حذف می
شوند،موتاسیون
های مفید جدید
به سبب فراوانی
کم در معرض اسیب
رانش ژنی قرار
دارند.
جدایی
محل سکونت
جدایی
محل سکونت
اشاره به موقعیتی
دارد که دو
گونه این
توانایی را دارند
که با هم زندگی
کنند اما در
محل های سکونت
متفاوتی زندگی
یا تولید مثل
می کنند.این
تفاوت در محل
زندگی دو گونه
را از هم جدا
نگه می
دارد.روش های
متفاوتی وجود
دارد که یک
محل سکونت
بتواند تقسیم
شود.
در این
مورد مثال های
گوناگونی
وجود دارد،
مثلا چیتاها
مکان عمومی
مشابهی را با leopard ها و شیرها
تقسیم می کنند
اما چیزی که
آنان را از دیگر
گربه سانان
جدا می کند این
است که چیتاها
عموما روز
فعال هستند در
حالی که شیرها
در شب فعالیت
خود را انجام
می دهند.

جدایی
زیستگاهی بین
دو گونه خواهری
neotropical costus :
سازش
به شرایط
اکولوژیکی
متفاوت به
عنوان یک مکانیسم
عمده گونه زایی
شناخته شده.در
این جا نقش
جدایی زیستگاهی
یک جفت گونه
نزدیک به هم
را که جزء علف
های neotropical
بودند بررسی
کردیم.
Costus allenis وCostus villossimus که میکرو
هابیت های
مجزایی را
اشغال کرده
بودند.
در panama مرکزی costus allenis زیستگاه
های بدون نور
خورشید (سایه
دار) و مرطوب
را که در
امتداد دره
قرار دارند ترجیح
می دهد در حالی
که c.villossimus در
جای خشک و
مکان های باز
در امتداد حاشیه
جنگل ساکن
است.
برای
تحقیق بزرگی
در مکانیسم های
سازش محلی:
1)توزیع
رسوب و رطوبت
خاک جمعیت های
طبیعی با هم
مقایسه شد.
2) یک پیوند
متقابل را برای
تعیین اینکه
شواهدی برای
سازش محلی
وجود دارد
،انجام دادیم.
3)آزمایشی
را برای تعیین
مقاومت به خشکی
گلخانه ای برای
دو نسل والد و
زاده های F1
انجام دادیم.
نتایج
و بحث:
یک
جدائی میکروجغرافیایی
بسیار قوی دربین
گونه ها در شیب
رسوبی دیده
شد.این توزیع
فضایی مجزا با
تفاوت در بارش
در طی فصل های
خشک مرتبط
است،با رسوب بیشتر
در منطقه شمالی
که بوسیله c.alleni
اشغال شده بود
نسبت به منطقه
جنوبی که بوسیله
c.villossimus
اشغال شده
بود. در
فصلی خشک در
سال 2007 ،درصد
متوسط رطوبت
خاک برای c.alleni
حدود دو برابر
بیش تر از c.villossimus
بود.
آزمایش
پیوند متقاطع
نشان داد c.alleni
قدرت بقاء بیشتری
را در زیستگاه
خودش دارد(77%) نسبت
به وقتی که در
زیستگاه c.villossimus است(35%) ،در
حالی که c.villossimus
نرخ رشد نسبی
بیشتری را در
زیستگاه خود
داشته (0.65±0.16)
نسبت به زیستگاه
c.alleni (-0.3±0.13) . آزمایش
خشکی گلخانه ای
نشان داد که c.alleni همه برگ
های سبز خود
را بعد از 140 روز
بی آبی از دست
داد در حالی
که نسل F1
هیبرید و c.villossimus
به ترتیب 3٪ و 17٪ از
برگ های سبز
خود را حفظ
کردند، که
نشان داد c.alleni
نسبت به خشکی
تحمل پایین تری
دارد که این
امر باعث نرخ
مرگ و میر بیشتر
c.alleni در
زیستگاه c.villossimus
می شود. همه شواهد پیشنهاد
می کند که میزان
خشکی با گونه
زایی زیستگاهی
c.alleni و c.villossimus
مرتبط است.
برای
تعیین اهمییت
این فاکتورهای
غیر زنده ما :
1)تحمل
نسبت به
خشکسالی را در
جمعییت هیبرید
نسل F2
تخمین زدیم.
2)انتخاب
بر روی این
صفت را در جمعیتی
آزمایشی با
ژنوتیپی
مشابه F2
که در حال رشد
در کشتزار
بودند اندازه
گرفتیم.
3)انتخاب
هایی را که به
واسطه عواملی
دیگر، به طور
عمده زیستگاه
های دو جمعییت
والدی را از
هم متفاوت می
سازد،اندازه
گرفتیم.(این زیستگاه
ها دسترسی به
نور داشتند و
دارای
جانوران گیاه
خوار بودند)
در
کل،این
مطالعه شواهدی
را نشان داد
که سازش محلی
و گونه زایی microhabitation در
مقوله گونه زایی
نقش دارد.
References:
1)wikipedia
2) Grace Fu-chun Chen and Douglas W. Schemske.
مقدمه:
همه ی
اورگانیسمها
زنده از یک جد
مشترک از35 میلیارد
سال پیش مشتق
شده اند.و جمع
شدن جهشهای
ژنتیکی باعث
بوجود آمدن این
تنوع زیستی که
در حال حاضر می
ببینیم شده
است. رد پای این
تکامل درژنوم
اورگانیسمهای
موجود باقی
مانده است و
با استفاده از
آنها می توانیم
روابط فیلوژنی
اورگانیسمها
وتغییرات ژنتیکی
را استنباط کنیم.
این روابط فیلوژنی
استنباطی بینش
وسیعی را در
رابطه بازمینه
های متنوع بیولوژی
تکاملی پیش روی
ما می گشاید .
تعریف
گونه زایی و
معرفی ژنهای
هومئوتیک:
گونه
زایی یک پرسه ی
تکاملی است که
یک گونه ی بیولوژی
جدید ایجاد می
کند.گونه زایی
بطور نرمال
بوسیله ی دریفت
ژنتیکی ویا
انتخاب طبیعی
رخ می دهد.
ژنهای
هومئوتیک در ایجاد
الگوی تکوین
در همه ی
اورکانیسمها
ضروری می
باشند،این
ژنها ی هومئوتیک
هستند که می
گویند هر قسمت
به چه چیز تکوین
یابد .زنهای
هومئوتیک در
همه ی حیوانات
بطور مشابه
ظاهر می
شوند.در سال 1980
ثابت شد که یک
توالی مشترک
حدود 180 جفت باز
در همه ی ژنهای
هو مئوتیک
وجود دارد، این
توالی مشترک
هموباکس نام
گرفت که تقریباًدر
همه ی یوکاریوتها
وجود دارد .
این
ژنها تنظیم بیان
دیگر ژنها و
کنترل پرسه های
تکوینی را بر
عهده دارند .
یک زیر
گروه مهم از
آنها که مخصوص
جانوران است
ژنهای هاکس می
باشند که
درالگوی بیان
اندامها ی
حرکتی بسیار
مهم می باشند .
زیر
گروه دیگر MADS-box است که
مخصوص گیاهان
است و در ایجاد
مدل ABC در
تکوین گل نقش
دارد .
HOX gene & speciation
بیان
ژنهای هاکس در
طی تکوین در
طول محورقدامی
خلفی موش
مشابه دروزوفیلا
است . مقایسه ی
سکانس ها ی این
ژن ها نشان
داده این ژنها
در بین دروزوفیلا
وموش حفظ شده
هستند .
بنابراین
ممکن است
تفاوتها در
طرح بدن مهره
دارن و حشرات
به تکامل شبکه
ها ی ژنی ،که
احتمالا بر هم
کنشهای تنظیمی
مشابهی دارند
ربط داده شود.

ژنهای
هاکس در مهره
دارن درایجاد
الگوی
اندامهای
حرکتی موثرند.
یک مثال از
آنها که در
جوانه ی اندام
حرکتی بیان می
شوند در زیر
نشان داده شده
است.

جهش
در یکی از
ژنهای هاکس
باعث ایجاد پای
نابجا در محل
ایجا آنتنها
شده است(شکل
سمت چپ)
ونمونه طبیعی
در شکل سمت
راست نشان
داده شده است.


MADS-box & spesiation
گل
اورگان تولید
مثلی در گیاهان
خشکی زی است و
شامل کاسبرگ
،گلبرگ ،پرچم
ومادگی می
باشد .سلولهای
زایشی هاپلوئید
ماده به وسیله
ی یک اسپورانژیوم
و دو اینتگیومنت
و سلولهای زایشی
نر توسط یک
اسپورانژیوم
پوشیده می
شوند.از طرف دیگر
سرخسها
وبازدانگان
اورگانهای
تولید مثلی
ساده تری نسبت
به
نهاندانگان
دارند وفاقد
کاسبرگ وگلبرگ
میباشند.اسپورانژیای
ماده در با
زدانگان فقط
با یک اینتگیومنت
پوشیده میشود
ودر سرخسها
فاقد اینتگیومنت
است .
تکوین
اورگان گل بوسیله
ی اعضای
خانواده ی
ژنهای MADS
تنظیم می شود
.این ژنها در
نهاندانگان
به 10 گروه تقسیم
می شوند .چه
نوع تغییری در
ژنهای MADSباعث
تکامل
اورگانهای
تولید مثلی در
دودمان گیاهان
گلدار شده
است؟
ژنهای
MADS ابتدا
در یک نوع
سرخس (Ceratopteris richardii) شناسایی
شدند .که فقط
از سه گروه ژنی
تشکیل شده است
که نسبت به گیاهان
دانه دار کمتر
است. الگوی بیان
5 ژن موجود در
هر 3 گروه ژن MADS سرخس
در طی اسپروفیت
وگامتوفیت
نشان داد که
بعضی ژنهای MADS فقط
در اورگان گل
بیان می
شونددر حالیکه
دیگر ژنها یMADS هم در
اورگانهای زایشی
وهم رویشی بیان
می شوند . اکثر
ژنهای MADS
سرخسها در هر
دو اورگان بیان
می شوند . بیان
در هر دو
اورگا ن ممکن
است نسبت به بیان
اختصاصی MADSدر
اورگان زایشی
ابتدایی تر
باشد .بنابراین
بعضی از ژنها ی
MADS که
اختصاصی
اورگانهای زایشی
بودند انتخاب
شدند .که
بعدها باعث ایجاد
واگرایی در بین
سرخسها و
نهاندانگان
شد .
آنالیز
ژنهای MADS
در بازدانگان(
که از نظر
اورگانهای زایشی
ما بین سرخس
وننهاندانگان
قرار گرفته
اند) در جریان
است.
محدودیت
بیان ژنهای MADSممکن
است بوسیله ی
تکامل دیگرژنهایی
که تنظیم بیان ژنهای MADS
را بر عهده
دارند توضیح
داده شود .ژن LEAFY یکی از
تنظیم کننده
های MADS هومئوتیک
متداول در Arabidopsis است .
نقشMADS در گیاهان
بدون گل هنوز
مبهم است
،خصوصیات
عملکردی ژن MADS در خزه
میتواند بینش
جدیدی در این
رابطه پیش روی
ما بگذارد.
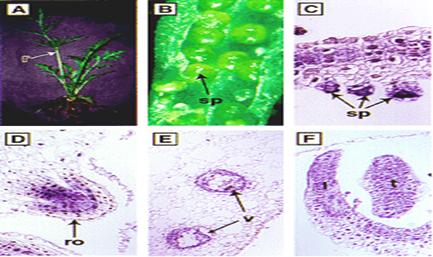
الگوی
بیان CMADS 1 RNA در
بافتهای
اسپروفیت بوسیله
ی دو رگه سازی
در جا . ژنهای MADS Ceratopteris richardii در هر
دو اورگان زایشی(C) و
رویشی (DEF) بیان شده
اند .(A
اسپروفیت C.richardiiیک
گیاه کامل با
برگهای رویشی
وزایشی.(B
اسپورانژیای
لخت دور از
محور اصلی
برگ.(C
مقطع طولی
اسپروفیل که
اسپورانژیا
نشان می دهد.(D
مقطع طولی از
ناحیه ی رشد
در نوک ریشه .(E
مقطع عرضی از
دمبرگ که vascular
bandle نشان
داده شده است. (Fمقطع طولی(l)و عرضی
(t)
از دو برگ رویشی
جوان .
هومولوژي
يا ساختارهاي
هومولوگ
ارگانهايي در
گونه هاي
متفاوت هستند
که داراي
ساختار مشابه
مي باشند و
منشا تکاملي
يکسان دارند و
از يک جد
مشترک بوجود
آمده اند . اما
ممکن است تفاوتهايي
در ظاهر و عمل
داشته باشند .
اين ساختار ها
مي توانند
عملي که در جد
مشترک داشته
اند حفظ کرده
و يا عملکرد
آنها بطور کلي
تغيير پيدا
کرده باشد . بنابراين
ساختارهاي
هومولوگ
لزوماً کار يکساني
انجام نمي
دهند. مانند
باله وال و
دست انسان.اين
دو عضو در
ظاهر متفاوت
هستند اما
ساختار
اسکلتي داخلي
آنها و ترتيب
قرارگيري
استخوان و
ماهيچه آنها
يکسان يا
بسيار شبيه
بهم مي باشند. هم چنين
بال پرنده و
بال خفاش نيز
از نظر اسکلت
بندي همولوگ
هستند.

ساختار
هاي همولوگ در
ارگانيزم هاي
تکامل يافته
تر در ظاهر
شباهتهاي
کمتري نشان مي
دهند ،اما با
اين حال
رديابي روابط
تکاملي بين آنها
امکان پذير
است.
در
مورد گياهان
نيز هر برگ در
گونه هاي
مختلف گياهان
شکل و عمل
متفاوتي دارد
با اين وجود
تمام آنها
ساختارهاي
هومولوگ هستند
، که از يک
ساختار
اجدادي مشترک
مشتق شده اند.

Molecular homology:
تاريخچه
تکاملي يک
گونه نشانه
هايي در DNA و
پروتئين هايي
که کد مي کند
بر جا مي گذارد.
دو
گونه که سکانس
DNA
مشابهي دارند
احتمالاً از
يک جد مشترک
مشتق شده اند.
(البته
سکانس هاي
مشابه هميشه
به دليل وجود
داشتن جد
مشترک نيستند
. سکانس هاي
کوتاه ممکن است
بصورت اتفاقي
بهم شبيه شوند
و يا به دليل اينکه
به يک پروتئين
خاص متصل مي
شوند شباهت داشته
باشند ، مثل
فاکتورهاي
رونويسي.)
تمام
ارگانيزم هاي
زنده براي به
رمز درآوردن اطلاعات
خود از 4 باز
آدنين ، تيمين
، سيتوزين و
گوانين
استفاده مي
کنند، و اين
واقعيت را خاطر
نشان مي کند
که تمام
ارگانيزم ها ي
زنده از يک جد
مشترک اوليه
مشتق شده اند.
مثال
ديگر در مورد
ژن pax-6 است که يک
ژن دخيل در
تکوين چشم
است. تمام
پستانداران ،
پرندگان ،
ماهي ها و حتي
حشرات داراي
اين ژن هستند.
ژن pax-6 عمل
مشابهي را در
تمام اين گونه
ها انجام مي دهد
و يک هومولوژي
محسوب مي شود.
اين ژن يک ژن
قديمي است که
در جد مشترک
تمام اين گونه
ها حضور داشته
است.



ساختارهاي
آنالوگ ساختارهايي
هستند که در
گونه هاي
متفاوت عملکرد
يکساني دارند
اما ساختار
آنها مشابه
نيست و از يک
جد مشترک نيز
مشتق نشده
اند.
شباهت
اين ساختارها
در پاسخ به
محيط زندگي مشابه
،روش زندگي
يکسان ، روش
جابجايي و
حرکت و يا
منبع غذايي
همانند و از
طريق co
evolution (تکامل
همگرا) ايجاد
مي شود.
مثال :
بال
حشرات و
پرندگان

بدن
عايق شده با
چربي و دوکي
شکل فوک (
پستاندار) و
پنگوئن (
پرنده ) که به
دليل شباهت
محيط زندگي
آنهاست.


Squirrel flying و Sugar glider
داراي شباهت
هاي بسياري
هستند. نظير :
چشم بزرگ ،
شکم سفيد و
پوسته نازک
بين دستها و
پاها که به
آنها در پريدن
از ارتفاعات کمک
مي کند.


اما Flying squirrel يک
پستاندار جفت
دار و Sugar
glider يک
پستاندار
کيسه دار است.
بنابراين
چشم بزرگ و
پوست بين دست
و پا در اين دو
گونه اندام
هاي آنالوگ هستند
که تحت فشار
انتخاب طبيعي
به دليل شرايط
زندگي يکسان
مثل پريدن از
ارتفاعات و
شکار در
تاريکي سازش
پيدا کرده
اند.

در
مورد گياهان
نيز، گياهان
صحرازي در
ظاهر شبيه به
هم و مثل
کاکتوس ها به
نظر مي رسند
در حالي که
شرايط خاص
محيط و سازش
با آن باعث
بوجود آمدن
اين خصوصيات
شده است.
خصوصياتي
مثل ساقه مومي
و پر از آب جهت
ذخيره آب ، و
تيغ هايي جهت
حفاظت از
جانوران
گياهخوار.


موانع
زیادی برای
تولید مثل بین
دو گونه مختلف
وجود دارد.هر
گونه ای نمایشهای
جفتگیری خاص
خود را دارد و یا
فصل تولید مثلی
مخصوص به
خود.بنابراین
افراد دو گونه
متفاوت شرایط
جفتگیری با یکدیگر
را ندارند و
موفق به
زادآوری نمی
شوند چون تخم
تشکیل نمی شود
و یا در مراحلی
از رشد از بین
می رود. دو سری
مکانیسم برای
جدایی تولید
مثلی پیشنهاد
می شود:
مکانیسم
جدایی تولید
مثلی prezygotic
که شامل جدایی
اکولوژیکی
،جدایی فصلی،
جدایی رفتاری
،جدایی مکانیکی
و جدایی گامتی
می باشد.
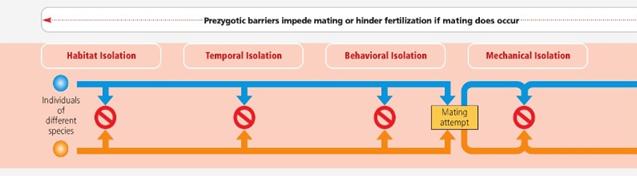
مکانیسم
جدایی تولید
مثلی postzygotic
شامل عدم زنده
ماندن هیبرید،عقیم
بودن هیبرید،
هیبرید ضعیفی
که نسل حاصل
از آن قابلیت
زیست پایینی
دارد.
جدایی
مکانیکی در نتیجه
ی عدم تناسب
شکل و اندازه ی
اندام جنسی در
دو گونه کاملا
سمپاتریک
اتفاق می افتد
وجفتگیری فیزیکی
را غیر ممکن می
سازد.همچنین
موانع مولکولی
نیز مانع
اتصال اسپرم
به تخمک و تشکیل
تخم می شوند.
عقیده بر این
است که جدایی
مکانیکی در
بند پایان به
ویژه در حشرات
و هزارپایان
اهمیت بیشتری
دارد.به عنوان
مثال در
سنجاقک; dragonfly نر
از یک جفت
قلاب برا
گرفتن ماده در
طول جفتگیری
استفاده می
کند ولی این
سنجاقک نر وقتی
که سعی می کند
با یک سنجاقک
ماده از گونه
ای دیگر جفتگیری
کند موفق نمی
شود زیرا
قلابهای جنسی
آن با فرم بدن
ماده تناسب
ندارد.در دو
گونه از حلزونها
نیز تلاش برای
جفتگیری
اتفاق می افتد
ولی تفاوتهای
مورفولوژیکی
مانع موفقیت
ان می شود.صدف
این دو گونه
از حلزون از
جنس Bradybaena در
جهات مختلف پیچ
خورده است.در یکی
از آنها مارپیچ
ها در خلاف
جهت گردش
عقربه های
ساعت و در دیگری
در جهت گردش
عقربه های
ساعت است.در
نتیجه منفذ
جنسی این
حلزونها همسو
نیستند و جفتگیری
کامل نمی شود.
در گیاهان
که اغلب بوسیله
ی حشرات گرده
افشانی می
شوند،آناتومی
گل غالبا با
گونه ی حشره ی
ویژه ای که
گرده های این
گونه از گیاه
را منتشر می
کند سازگار
شده است.جدایی
مکانیکی بیشتر
در گیاهان
گلدار اتفاق می
افتد .گلهای
گل مریم سیاهsalvia officinalis با
گل مریم سفید
از لحاظ
ساختاری
متفاوت است و
توسط گونه های
متفاوتی از
حشرات گرده
افشانی می
شوند.در این
مثال هر گونه
از حشره تنها
گرده ی یک
گونه از گل مریم
را منتشر می
کند بنابرین
زادآوری بین این
دو گونه اتفاق
نمی افتد.

یک
حالت جالب اینکه
;
در دو گونه از
مویزکlousewort
که هر دو توسط
گونه ی زنبور
عسل Bumblebee
گرده افشانی می
شوند جدایی
مکانیکی چنین
اتفاق می افتد
که گرده های
هر گونه از مویزک
توسط قسمت خاصی
از بدن حشره
منتقل می
شوند.
موتاسیون
تغییر توالی
نوکلئوتیدی
درارگانسیم
ها است و
موجودی که
فنوتیب جدید
را نشان می
دهد جهش یافته
نام دارد .در
اواخر قرن
نوزدهم قبل از
کشف قوانین
مندل Devries این
تغییرات را
جهش نامید.
این
تغییرات شامل
تغییر در
تعداد
کروموزوم ها(aneuploidy و euploidy) و
تغییردر
ساختمان
کروموزوم ها
است. ولی کلمه
جهش فقط جهت
تغییرات ژن به
کار می رود
جهش ها یا در
سلول های
سوماتیکی است یا
در سلول های
جنسی جهش هایی
که در سلول های
جنسی است قابل
توارث است
جهش
ها در اثر
اشعه ها است
که هر چه طول
موج کوتاهتر
باشد انرژی آن
بیشتر و قدرت
نفوذ آن نیز بیشتر
است
مثل
اشعه x
ماورابنفش در
اثر مواد شیمیایی
مثل آلکالات و
گاز خردل ویا
تصادفی است
Transition
Point mutation
← یک n.t جایش با
نوکلئوتید دیگر
عوض می شود که 2
دسته
Transiton
← یک
پورین جایگاهش
با پورین دیگر
عوض می شود (A↔G)
Transversion
یا یک
پیریمیرین با
پیریمیگ دیگر
(T↔C).
←
Transversion یک پورین
با یک پیریمیدین
جایگاهش عوض می
شود و بالعکس (S/T ↔A/C).
جهش
های نقطه ای
که در کد شدن
پروتئین ها روی
می دهد براساس
کدن ترجمه
شونده به 3
دسته تقسیم می
شود .
Silent mutation
← کد ایجاد
شده همان aa اصلی را
کد می کند
Missense mutation ←
اسید آمینه جدیدی
کد می شود
← Nonsense
mutation کدن پایان
ایجاد شده و
آمینو اسیدی
تولید نمی شود
Incertions mutation ←
اضافه شدن چند
نوکلئوتید به DNA که اگر ntها
اضافه شده 3 یا
مضربی از 3
باشد Frame shift به هم نمی
خورد.
Deletion mutation ← یک
یا چند n.tازDNA
حذف شده است
که می تواند reading frameرا
تغییر دهد یا
خیر.
(در
صورت تغییرreading frame
ناهنجاری ها
بسیار شدید
خواهد بود و
موجب مرگ
ارگانیسم می
شود )
جهش
ها در مقیاس
بزرگ در
ساختار
کروموزمی است
.
Amplification
← 2
برابر شدن یک
ژن
← Deletation حذف یک
بخش کروموزومی
Fusion gene
←پهلوی
هم قرار گرفتن
2بخش جداگانه
ازDNA که شامل :
← Chromosomal
translocationپهلوی
هم کنار قرار
گرفتن 2 بخش
ژنتیکی از
کروموزوم های
غیر همولوگ
Interstitial delletion← حذف
یک بخش ازDNA
که بخشی از
کروموزوم است
← Choromosomal
inversionمعکوس
شدن یک بخش از
کروموزوم
Loss of heterozygosity
← از
دست دادن یک
الکل در یک
کروموزوم
(البته
تقسیم بندی دیگری
نیز برای
موتاسیون ها
داریم که بر
اساس عملکرد و
تاثیر آن ها ,
اثرات آن ها,
وراثت, اثر روی
توالی پروتین
ها وبر اساس
شایستگی.)
بر
اساس عملکرد:
Loss of function mutation
←
محصولات ژن
عملکرد ی
ندارد یا
عملکرد ناقص
است کAmorphic mution
نیز نامیده می
شود
Gain of function mutation ←
ساختار محصول
ژن عوض شده غیر
نرمال دارد
Dominant negative mutation ←
antimorphic mutation
Letal mutation
←جهش
های کشنده که
ارگانسیم
ناقل ژن می میرد
Back mutation
← یکpoint mutation است
که یک توالی
جهش یافته را
به توالی اصلی
و نرمال تبدیل
می کند
بر
اساس وراثت
← heterozygous
mutation جهش در یک
الل
← homozygous
mutation در هر دو
الل مادری و
پدری
بر
اساس وراثت روی
توالی پروتئین
ها
← Frame shift mutation در اثر insertion یاdeletation یا که
چهارچوب
خواندن و
ترجمه می شود
← Missense یک نوعpoint mutation که
باعث توقف بیشتر
می شود
← Neutral mutationجهش های
خنثی
← Nounsense mutationیک نوعpoint mutation است
کهstop codon ایجاد
میشود
← Silent mutationهر دو
رمز یک اسید
آمینه را کد می
کنند
بر
اساس شایستگی :
1.
Harmful mutation
2.
Beneficial mutation
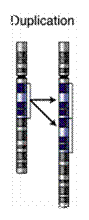
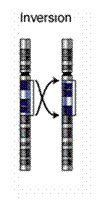


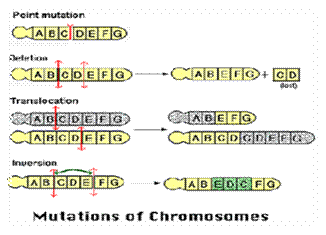
انتخاب
طبیعی یکی از
پایههای بیولوژی
مدرن میباشد.
اصطلاحی که
اولینبار
توسط چارلز
داروین تعریف
شد.

انتخاب
طبیعی چیست ؟
و چگونه
به تغییرات
تکاملی منتهی
می شود؟
انتخاب طبیعی
تفاوت آشکار
در بقا وتولیدمثل،
میان ژنوتیپها
و یا حتی ژنهای
مختلف میباشد.
(ژنوتیپ: گروهی
از ارگانیسمها
که مواد ژنتیکی
معینی را دارا
میباشند.) فرایندی
است که بوسیله
آن گونهها با
محیطشان سازش
مییابند.
انتخاب طبیعی
وقتی به تغییرات
تکاملی منتهی
میشود که میزان
تولیدمثل یا
بقای جمعیتی
با صفات معین،
از سایر افراد
جمعیت بیشتر
باشد و آنها
بتوانند صفات
ژنتیکی خود را
به نسلهای
بعد منتقل
کنند. انتخاب
طبیعی حاصل
تنوع، تولیدمثل
افتراقی و
وراثت میباشد.

آیا
انتخاب طبیعی
تنها مکانیسم
تکامل است؟
قطعا
نه، پیشنیاز
برای انتخاب
طبیعی، تنوع و
تفاوت ژنتیکی
قابل توارث میباشد.
تنوع ژنتیکی
از جهش، نوترکیبی
و تغییر در
کاریوتیپ
حاصل میشود.
درواقع تکامل
بدون تنوع ژنتیکی
نمیتواند ایجاد
شود. ولی بهمحض
ایجاد تنوع
ژنتیکی دو
احتمال وجود
دارد:
genetic
drift :در این
حالت تفاوتی میان
ژنوتیپها و یا
ژنهای مختلف
در اثرشان بر
بقا یا تولیدمثل
وجود ندارد. تنها تغییرات
تصادفی یک فرد
در مقابل سایرین
در یک جمعیت
وجود دارد.natural
selection: که تغییرات
قابل پیشگویی
در نسبت یک ژن یا
ژنوتیپ به دیگری
میباشد و پایدارتر
است.
آیا
انتخاب طبیعی
به تولید گونه
جدید منتهی میشود؟
چگونه؟
در
برخی موارد.
بخش عمدهای
از تکامل،
بدون تشکیل
گونه جدید،
توسط انتخاب
طبیعی اتفاق میافتد.
انتخاب طبیعی
تنها فرایند
سازش درون
گونهها است.
در برخی شرایط
انتخاب طبیعی
در تشکیل گونه
جدید نقش
دارد. برای
مثال تشکیل
دو گونه
شامپانزههای
امروزی و جنس
انسان از
دودمان پریمات
اجدادی.
تصورات
غلط درمورد
انتخاب طبیعی
چیست؟
1.
انتخاب طبیعی
باعث بقای شایستهترین
(fittest) میشود.
اینطور نیست.
زیرا گاهی
اوقات نوع شایستهتر
وجود ندارد.
ممکن است
انواع متفاوتی
که بهطور
مساوی شایسته
میباشند وجود
داشته باشد.
شاید آنها به
جنبههای
مختلف محیط سازش
یافتهباشند.
دراین صورت هیچ
یک با دیگری
جایگزین نمیشود.
2.
انتخاب طبیعی
هدفمند عمل میکند
و درجهت برطرفکردن
نیاز ارگانیسم
پیش میرود. خیر،
انتخاب طبیعی
تنها میان
تنوعی که در
جمعیت وجود
دارد انتخاب میکند.
درطرف مقابل
گاهی اوقات،
انتخاب طبیعی
بهعنوان فرایندی
تصادفی تعبیر
میشود. تنوع
ژنتیکی که
درنتیجه جهش
رخ میدهد،
تصادفی است ولی
انتخاب طبیعی
بهصورت غیرتصادفی
بر تنوع ژنتیکی
عمل میکند.

اتخاب
طبیعی بر اساس
فرکانس آلل،
به چند نوع
تقسیم میشود؟
1. :Directional Selection زمانیکه
بدلیل برتری یک
آلل نسبت به
سایر آللها،
فرکانس آن آلل
در جمعیت افزایش
مییابد و
درنتیجه منحنی
به سمت راست یا
چپ شیفت میکند.
به بیان دیگر
انتخاب طبیعی
بهنفع افرادی
از جمعیت میباشد
که در یک کران
توزیع صفت
قرار دارند. بهعنوان
مثال مقاومت
باکتریها به
آنتیبیوتیک.
فسیلهایی که
نحوه تکامل
اسب را بیان میکند
شاهد خوبی برای
این نوع
انتخاب طبیعی
میباشد.
2. Stabilizing
Selection (Purifying): شایعترین
مکانیسم
انتخاب طبیعی
است. در این نوع،
انتخاب طبیعی
بهنفع فنوتیپ
حدوسط میباشد
. درواقع
همراه با ثبات
جمعیت در یک
صفت معین،
تنوع ژنتیکی
کاهش مییابد.
بهعنوان مثال
Siberian Husky نوعی
سگ است که
عضلات سینه و
پای بسیار قوی
دارند تا
بتوانند بهراحتی
در برف حرکت
کنند. اگر این
سگها عضلات
حجیم داشته
باشند، در برف
فرو میروند و
با عضلات ضعیف
هم نمیتوانند
سورتمهها را
بکشند. بنابراین
انتخاب
طبیعی برای
اندازه عضله، یک
میانگین (norm) را
انتخاب کرده
است.
3. Disruptive
Selection: در این
نوع انتخاب طبیعی
دو فنوتیپ
مختلف،
همزمان در محیط
وجود دارند و
درنتیجه در
منحنی توزیع
صفت، دو پیک
مشاهده میشود.
بهعنوان مثال
فنچهای افریقا
که یا نوک
بلند یا نوک
کوتاه دارند و
اندازه حدوسط یافت
نمیشود.

{kind=link}
parapatric
و parapatry عباراتی
از biogeograghy
هستند و به
ارگانیسم هایی
برمیگردند که
محدوده آنها
باهم overlap
ندارند اما
مجاور یکدیگر
هستند.
Parapatric speciation یکی از فرم های
گونه زایی است
که به دلیل
تنوع در
فرکانس جفت گیری
یک جمعیت در یک
ناحیه جغرافیایی
با گرادیان محیطی
خاص ایجاد میشود.در
گونه زایی
پاراپاتریک هیچ
سد خارجی خاصی
برای جریان ژنی
وجود
ندارد.جمعیت
ادامه پیدا میکند
اما افراد جمعیت
به طور تصادفی
جفت گیری نمیکنند. افراد
جمعیت بیشتر
تمایل دارند
با همسایگان
جغرافیایی
خود جفت گیری
کنند تا با
افرادی در بخش
های مختلف
محدوده جمعیت.
Divergence به
دلیل کاهش جریان
ژنی در جمعیت
و اختلاف فشار
انتخابی در
محدوده جمعیت
اتفاق میافتد.
گونه
زایی
پاراپاتریک
را در گیاه
گونهAnthoxanthum odoratum میتوان دید.
برخی
از این گیاهان
نزدیک معدن
زندگی میکنند
که در این ناحیه
خاک با فلزات
سنگین آلوده میشود.
گیاهان اطراف
معادن تحت
انتخاب طبیعی
برای ژنوتیپ
هایی که به عناصر
سنگین مقاوم
هستند قرار میگیرند.
در حالی که گیاهانی
که دور از این
منطقه هستند
تحت این
انتخاب طبیعی
برای این صفت
قرار نمیگیرند.
دو نوع گیاه
آنقدر به هم
نزدیک هستند
که گیاهان
مقاوم و غیرمقاوم
پتانسیل لقاح
با یکدیگر را
داشته باشند.
بنابراین دو
نوع گیاه تکامل
مختلفی را طی
میکنند که
منجر به زمان
گلدهی متفاوت
در آنها میشود.تفاوت
در زمان گلدهی
باعث جدایی
تولید مثلی
دراین دو گروه
میشود ودر نتیجه
این تغییر اولین
مرحله برای
قطع جریان ژنی
بین دو گروه میشود.

فنتیک
واژه
فنتیک به نام
تاکسونومی
عددی یا تاکسی
متری شناخته
شده است و
کوششی است که
برای طبقه بندی
ارگانیسم ها
بر پایه
تشابهات کلی
،به طور معمول
درمورفولوژی یا
ویژگیهای
قابل مشاهده
،علیرغم فیلوژنی
یا رابطه
تکاملی شان
انجام میشود.
فنتیک
به طوروسیعی
بتوسط کلادیستیک،
برای تحقیق
ارتباطات
تکاملی بین
گونه ها جای
گزین شده است.
متدهای
اصلی فنتیک
،مثل پیوستگی-همسایگی
،راهشان را به
کلادیستیک به
عنوان تخمین
مستدّل فیلوژنی
،وقتی متدهای
پیشرفته تر
مثل (استنباط
بایسین)از
لحاظ محاسباتی
گران هستند،پیدا
کرده اند.
تکنیک
های فنتیک
شامل فرمهای
مختلفی از
دسته بندی و
گروه بندی می
باشد که راههای
پیچیده کاستن
تغییرپذیری
نمایش داده
شده توسط
ارگانیسم ها
به یک سطح
قابل مدیریت
است
در
واقع این به
معنی اندازه گیری
دوجین تغییرات
و بعد آماده
کردن آنها به
عنوان 2یا
3منحنی دارای
بعد،میباشد.
اغلب
چالشهای تکنیکی
در فنتیک حول
وحوش توازن
فقدان
اطلاعات ،به
طوری که کاهش،
برخلاف آسانی
تفسیر نمونه
های حاصله
باشد، دورمیزند.

تفاوت
فنتیک و کلادیستیک
آنالیزهای
فنتیک ،بین ویژگیهای
پلسیومورفی
که از یک جد
مشترک به ارث
میرسند ( که از
لحاظ فیلوژنتیکی
فاقد اطلاعات
هستند ) و ویژگیهای
آپومورفی (که یک
یا چندین
دودمان جدید
رااستنتاج میکند
) تمیز نمیدهد. بنابراین
آنالیزهای
فنتیکی ، به
توسط تکامل
همگرا وتشعشع
سازشی مشمول
گمراهی شده
است. یک خطای
نوعی رخ دهنده
در آنالیز فنتیک،طبقات
تکاملی پایه ای
است که بسیاری
از پلسیومورفی
ها را در مقایسه
با دودمانهای
پیشرفته که به
نظر می آید
مونو فیلتیک
باشند، حفظ میکند. مثال
پرندگان آوازی
را ملاحظه کنید.
اینها میتوانند
به دو گروه
تقسیم شوند. ( کورویدا )
که خصوصیات
اجدادی خود را
در فنوتیپ
وژنوتیپ حفظ
کرده اند و ( پسریدا )
که ویژگیهای
جدید بیشتری
دارند. امّا
فقط دوّمی یک
گروهی از نزدیکترین
خویشاوندان
است، اولی به
طور مستقل
پرجمعیت است و
دودمانهای
باستانی آن به
طوری فاصله
دار به هم
وابسته هستند. دریک
آنالیز فنتیکی
،درجه وسیعی
از تشابهات کلی
در بین کورویدا
پیدا شده که
باعث شده آنها
به نظر مونوفیلتیک
برسند. امّا ویژگیهای
مشترک بین
آنها دراجداد
تمام پرندگان
آوازی هنوز
حضور دارد.این
فقدان ویژگیهای
اجدادی نسبت
به حضور
آنهاست که این
موضوع را که
کدام پرندگان
آوازی،
وابستگی نزدیکتری
به همدیگرنسبت
به دیگر
پرندگان آوازی
دارند، معنی
دارمی کند. امّا دو
متدولوژی، نیاز
نداشت که به
طور دو طرفه
انحصاری شود.
به طورکلی تشخیص
داده شده که
فنتیک
امروزه،اطلاعات
غیرقابل اطمینان
بیشتری را
درباره
ارتباطات
تکاملی مابین
تاکسونها،برای
اینکه یک متد
قابل اتکا باقی
بماند،فراهم
میکند. هیچ دلیلی نیست
که چرا گونه
های مثال زده
شدهٔ شناسایی شده
با استفاده از
فنتیک ،نمیتواند
در معرض آنالیز
کلادیستیک
قرار گیرد؟ (
برای مشخص
کردن وابستگی
بین آنها ) متدهای فنتیک
،میتوانند از
کلادیستیک
وقتی که تنها
وضوح
تاکسونهای
وابسته مهم
است،بالاتر و
ممتازتر
باشند . در صورتی
که نیازهای
محاسباتی پایین
باشند. به عبارت دیگر،
هرگاه
اطلاعات راجع
به تاریخ
تکاملی
تاکسونها
،برای یک
مطالعه نیاز
است، یک محقق
امروزی سعی در
آنالیز با
استفاده از
متدهای کلادیستیک
دارد.
فنتیک
امروزی
به
طوری که از
اخبار برمی آید،
مناظرات داغ
وسیعی بین فنتیک
ها و کلادیستیک
ها ، به این
عنوان که هر
دو متد که در
آغاز برای حل
ارتباطات
تکاملی مورد
هدف بودند،
وجود داشت. شاید
night-water-markدر فنتیک
،مطالعات هیبریداسیون
DNA-DNA،بوسیله Burt L More,John E Ahlquist,Charles G Sibley
بود که ازآنها
تاکسونومی
پرندگان Sibley-Ahlquist
نتیجه شد. به طور خیلی
بحث آمیز در این
زمان، برخی از
یافته های
آن،به عنوان
مثل (
گالونسرا )
حمایت میشوند،
درحالیکه
مابقی مثل ( سیکونی
فورمها یا
کورویدا ) رد
شده اند. بسیاری از سیستماتیسیست
ها، استفاده
از متد فنتیک،
بویژه درسطح
آدرس دادن
گونه ها، را
ادامه میدهند. وقتی
که هدف نهایی
تاکسونومی
شامل یافتن «
درخت زندگی »
است، سیرتکاملی
مرتبط کننده
همه گونه ها-
در کارهای
صحرایی کسی را
نیاز دارد که
قادر به جدا
کردن یک
تاکسون از دیگر
تاکسونها
باشد. به
علاوه، بسیاری
از تکنیک های
توسعه یافته
توسط
تاکسونومیست
های فنتیک،
توسط انجمن
اکولوژیست ها
، در نتیجهٔٔ نیاز
مشابه به
ارتباط
برقرار کردن
با دسته وسیعی
از اطلاعات،
توسعه یافته و
پذیرفته شده
است.
منبع
:
www.wikipedia.com
پلي
پلوييدي
پلي
پلوييدي
حالتي در سلول
ها يا موجودات
است كه هر
سلول بيش ازدو
سري
ازكروموزوم
هاي همولوگ را
داشته باشد.
در حاليكه
بيشتر
موجودات حالت
ديپلوييدي
دارند و در هر
سلول دو سري
ازكروموزوم
هاي همولوگ
وجود دارد.
انواع پلي
پلوييدي ها بر
اساس
تعدادسري هاي
كروموزومي
نشان داده مي
شوند.
تري
پلوييدي (سه
سري كروموزوم 3X)
براي مثال در
نژاد جانوران
كندرو
تترا
پلوييدي (
چهار سري
كروموزوم4X ) براي مثال
در
آزاد ماهي ها
پنتا
پلوييدي (پنج سري
كروموزوم5X)
هگزا
پلوييدي( شش
سري كروموزوم6X
) براي
مثال درگندم و ميوه
كيوي
اكتا
پلوييدي ( هشت
سري كروموزوم
8X) براي
مثال درAcipenser گونه
هايي از سگ
ماهي
دكا
پلوييدي ( ده
سري كروموزوم10X) براي مثال
دربرخي
توت فرنگي ها
دودكا
پلوييدي(
دوازده سري
كروموزوم12X) براي مثال
درگياه
Celosia argentea
در
حيطه پلي
پلوييدي ها ما
بايد تفاوت
بين اتو پلي
پلوييدي و آلو
پلي پلوييدي
را بدانيم.
اتو پلي
پلوييدي از سري
هاي كروموزوم
هايي تشكيل
شده است كه
ازيك گونه
منشا گرفته
اند. موز و سيب
ميتوانند در حالت
اتوتري
پلوييدي يافت
شوند. آلو پلي
پلوييدها از
سري هاي
كروموزوم
هايي تشكيل
شده است كه
ازدو يا چند
گونه متفاوت
منشا گرفته اند.گندم
هاي زراعي
امروزي مثالي
ازيك آلو پلي
پلوييد است كه
از داراي شش
سري كروموزوم
است كه چهار
سري آن از
گندم (Triticum turgidum) و دو سري از
چاودار (Secale cereale) منشا گرفته
است. آلو پلي
پلوييدها فقط
ازگونه هاي
خيلي نزديك به
هم تشكيل مي
شوند و
كروموزوم هاي
متفاوت بصورت
كامل با هم
همولوگ نمي
شوند.
درحاليكه در
اتو پلي
پلوييدها
همولوگ شدن كامل
است.
پلي
پلوييدي در
گياهان شايع
است و تخمين
زده ميشود كه 30
تا 80 درصد گونه
هاي گياهان
زنده پلي پلوييد
هستند. پلي
پلوييدي
واقعي در
انسان بندرت
اتفاق مي افتد
هر چند در
بعضي بافتها
مانند كبد
وجود دارد. در
مورد جانوران
حالت پلي
پلوييدي
بيشتر در
جانوران پست
مانند كرم هاي
پهن و زالو ها
و ميگوي آب هاي
شور شايع است.
جانوران پلي
پلوييد اغلب
عقيم هستند و
بوسيله
پارتنوژنز
توليد مثل مي
كنند.
مامولكهاي
پلي پلوييد
نيززيادند و
بوسيله
پارتنوژنز
توليد مثل مي
كنند. سمندر
هاي زيرزميني
پلي پلوييد كه
اغلب تري
پلوييد هستند
همه شان ماده
هستند و
بوسيله
كليپتوژنز
توليد مثل مي
كنند. در اين
حالت اسپرم
بدون محتواي
ژني ازفرد
نربا تخمك را
بارور ميكند
تا موجود جديد
را توليدكند.
در حاليكه
سلول هاي كبدي
پستانداران
پلي پلوييد
هستند ولي
پستاندار پلي
پلوييد بندرت
شناخته شده
است چون اغلب
در حالت جنيني
مي ميرند. يك
استثناي
شناخته شده
ازاين قانون
يك جونده
octodontid در
مناطق بياباني
بد آب و هواي
آرژانتين
بنام موش
ويسكاچاي قرمزاست
(Tympanoctomys barrerae) .
اين جونده يك
موش نيست اما
خويشاوند خوك
هندي و
chinchillasاست. در
حالت 2n داراي
102كروموزوم
است و سلول
هايش حدودا دو
برابر اندازه
نرمال است.
خويشاوند
زنده نزديك به
اين موجود، موش
ويسكاچاي
آندان (
Octomys mimax)
كه 2n=56 است.
حدس زده مي
شود كه يك
موجود قديمي
شبيه Octomys حالت
تتراپلوييد 4n=112
را در اثر
دوپليكاسيون
كروموزوم
هايش توليد كرده
است . و در
نهايت اين
موجود در اثر
مكانيسم هاي
مانند
جابجاييهاي
كروموزومي
موجود مستقل
امروزي را
بوجود آورده
است.
جدایی
تولید مثلی
اساس گونه زایی
و فاکتورهای بیو
لوژیکی وجود
دارند که از Interbreeding بین
گونه های جدید
حاصله جلوگیری
می کنند تا
فرزندانی
بوجود بیاورند
که زایا باشند
و سدهای تولید
مثلی مختلفی
که خزانه های
ژنی را از
گونه ها جدا می
کند و بصورت Prezygotic Isolationیا Postzygotic Isolation
طبقه بندی می
کنند بسته به
اینکه عملکرد
انها قبل یا
بعد از تشکیل
زیگوت باشد.
Postzygotic Isolation:
بعد
از اینکه زیگوت
های هیبرید
تشکیل میشوند postzygotic isolation
اتفاق میافتد
زیگوت ممکن است
خیلی زود بمیرد
یا ضعیف باشد یا
اینکه عقیم
شود این مکانیسم
بعد از لقاح
رخ میدهد در
حالیکه جنین
در حال تکوین
است ومکانیسمهای
ان شامل مرگ و
میر زیگوتی
،زنده نماندن
هیبریدوعقیمی
است.گاهی
اوقات لقاح بین
گو نه های
مختلف رخ میدهد
اما جنین هیبرید
ضعیف است و میمیرد
در برخی موارد
هیبریدها قوی
هستند اما عقیم
اند. اگر لقاح
اتفاق بیفتد
چندین سد برای
هیبرید وجود
دارد.اولی
گامتی
است:گامتها
بطور موفقی با
هم ترکیب میشوند
اما بلافاصله
قبل از اینکه
هر تقسیمی
بتواند رخ دهد
میمیرند.دومی
زیگوتی است:زیگوت
تشکیل میشود
اما سریع میمیرد.سومی
جنینی یا لاروی
است:جنین هیبرید
بطور خودبخودی
از بین می
رود.چهارمی
عدم حیات هیبرید
است:زاده بدنیا
میاید اما
نامتناسب است.
سریعا با فشار
محیطی از پا
درمیاید و میمیرد.پنجمی
عقیمی هیبرید
است که هیچ
زاده ای نمیتواند
تولید کند
وژنهای ان از
هر دو گروه والد
جداست.در نهایت
هیبرید هایی
که زاده تولید
می کنند بطور
بالقوه ای
اخلاف عقیم
تولیدمی کنند
از بین رفتن
نسل F2 از
چرخش دوباره
مسیر ژنتیکی
در گونه های
مشخص جلوگیری
می کند. در بین
زاده های هیبریدی
که که بطور
نرمال تکوین میابند
و به بلوغ جنسی
میرسند
اکثرشان عقیم
هستند به این
معنی که انها
گامت های زنده
تولید نمی
کنند برای
مثال قاطر نتیجهای
از جفتگیری یک
اسب و الاغ
است انها بدنیا
می ایند و
بصورت حیوان
با لغ نرمال
تکوین می یا
بند اما
انهانمی
توانند زاده ای
از خودشان تولید
کنند.برخلاف
قاطرها اکثر
که در مرحله هیبریدی
زنده می مانند
تا بدنیا بیایند
سالم نیستند
اکثرشان قبل
از اینکه به
سن تولیدمثلی
برسند می میرند.
تاثیر
جمعیتهای
اجدادی روی
جدایی Postzigotic
در گونه زایی
الوپاتریک :
این
جدایی بواسطه
تجمعی از جانشینی
ها(اللهای حذفی
بالقوه در هیبریدها)در
جمعیتهایی که
از نظر جغرافیایی
جدا شده انداین
اللهای حذفی
بالقوه ممکن
است در جمعیتهای
اجدادی قبل از
جدایی جغرافیایی
حفظ شده باشند
نتایج نشان
داده که اللهای
دجنره شده در یک
جمعیت اجدادی
که در تعادل
قابل ملاحظه ای
ثابت باقی
مانده اند روی
جدایی Postzigotic
بین جمعیتهای
الو پاتریک
زادگان تاثیر
دارد سپس کاهش
شدید در
اندازه جمعیت(برای
مثال کمتر از
10نفر) برای
تکامل سریعPostzygotic isolation
لازم نیست وقتی
یک تفاوت زیادی
در شمار اللها
ی جمع اوری
شده بین احداد
و جمعیت
زادگان وجود
دارد گونه زایی
سریع رخ می
دهد ودر یکجمعیت
اجدادی با اندازه
جمعیتی موثر
کوچک که برای
دوره طولانی
حفظ شده جدایی
Postzigotic
بندرت تکامل می
یابد اگر
موتاسیونهایی
ایجاد شوند که
عملکرد اللهای
دجنره شده را
به حالت اول
برگردانند.نوترتیبی
کروموزومی میتواند
جدایی تولیدمثلی
را بوسیله
کاهش نوترکیبی
در طول یک
قطعه بزرگ از
ژنوم پیش
ببرد.اگر جدایی
بوسیله تجمع
مجموعه هایی
از اللهای
سازگار درون
گونه ای در Parapatry اما
ناسازگار در میان
گونه ها تولید
میشوند.نوترتیبیهای
کروموزومی
احتمالا نسبت
به سدهای ژنتیکی
بدون تغییر
کروموزومی ببیشتر
اتفاق می افتد
جدایی Postzygotic اغلب بواسطه
تجمع چنین
ناسازگاریهایی
است.
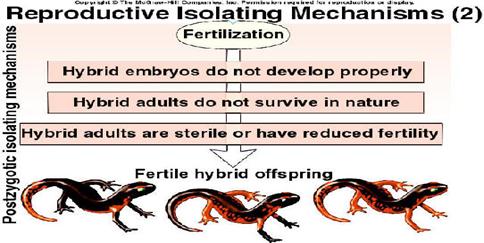 تr
تr
RAEFERENCE:
1)Takehiko I. Hayashi;
Masakado KawataImpact of ancestral populations on
postzygotic isolation in allopatric speciation Apr. 1, 2006 .
2)wikipedia
Cladistics :يك
روش آناليز
روابط تكاملي
بين گروههاي
ميباشد تا
درخت
خانوادگي
آنها ايجاد
شودواصل براين
است كه
موجودات بايد
بر طبق روابط
تكاملي شان
طبقه بندي
شوند. و راهي
براي يافتن
اين روابط اين
است كه صفات
اوليه و صفات
اشتقاق يافته
مشخص شوند.
Primitive characters: صفات
اوليه صفاتي
از گياه يا
جانور ميباشند
كه همه اعضاي
گروه داراي آن
ميباشند. به
عنوان مثال
وجود مو براي
يك صفت اوليه
است كه آنها
اين صفت را از
جد مشترك خود
به ارث برده
اند. (
پستانداران
اوليه يا
پستاندار
شبيه خزنده). و
مثال ديگر
داشتن غدد
پستاني و مو
در انسان و سگ
و گربه و
والها به
عنوان يك صفت
مشترك ميباشد.
و
مثال ديگر backbone يك
صفت اجدادي در
تمام مهرهداران
ميباشد.

اين
صفات در
آناليز روابط
تكاملي
موجودات با يك
گروه خاص
استفاده نميشوند.براي
رسم درخت
تكاملي در
پستانداران
مفيد نيست تا
يادداشت كنيم
كه آنها داراي
4 اندام حركتي
هستند. صفات
اوليه مشترك
در همه اعضاي
يك گروه symplesiomorphy ناميده
ميشودو در جد
مشترك قبلي تر
و قديمي تر
وجود دارد.
اين
صفات نميتواند
براي
كلاديستيك
كمك كننده
باشد و نبايد
براي رسم
كلادوگرام از
اين صفت
استفاده كرد ولي
براي مقايسه
دو گروه ميتوان
از آن استفاده
كرد. به عنوان
مثال داشتن 4 پا
در اسبها كه
در اسبهاي
قديمي تا
اكنون يك صفت
مشترك است
صفات
پيشرفتهاي
هستند كه تنها
در برخي از
اعضاي گروه
ديده ميشوند.:Derived characters
كلاديستيك
بر اين اصل
استوار است كه
ظهور صفات
اشتقاق يافته
نشانه اي از
روابط تكاملي
بين موجودات
است .
به
عنوان مثال در
دوزيستان و
لاك پشتها و
مارمولكها و
مارها و
كروكوديلها و
پستانداران و
پرندگان 4
اندام حركتي
دارند. در
مارههاي كنوني
4 اندام حركتي
را بطور واضح
نميبينيم
ولي در مطالعه
در شواهد
فسيلي ميبينيم
كه در مارهاي
قديمي اين 4
اندام مشخص
بوده
است.داشتن 4
اندام حركتي
يك صفت اشتقاق
يافته است.

يك
صفت اشتقاق
يافته ديگر
براي برخي
پستانداران
ممكن است كاهش
دم در آنها
باشد كه در
برخي از آنها
اتفاق افتاده
است مثلا
درانسانها و ميمونهاي
بزرگ. فرض بر
اين است كه از
دست دادن دم
در جد مشترك
انسانها و
ميمونها رخ
داده و آنها
اين صفت را از
جد مشترك خود
به ارث برده
اند. بنابراين
پستانداران
به دو گروه
داراي دم و
بدون دم تقسيم
ميشوندو
بوسيله دو
شاخه در
دياگرام
تكاملي نشان
داده ميشوند.صفات
اشتقاق يافته
در زبان
كلاديستيك آپومرفيك
ناميده ميشوند.
اگر
اين صفات تنها
متعلق به يك
گروه باشند به
آنautapomorphic مي
گويند. مانند
راه رفتن بر
روي دو پا كه
مخصوص
انسانها ميباشد
وautapomorphic
ناميده ميشودو
در اين صفت ما
با ميمونها
مشترك نيستيم.
:Synapomorphy يك
صفت اشتقاق
يافته است كه
در همه اعضاي
گروه ديده ميشود
مثلا از دست
دادن دم يك synapomorphy در
ميمونها و
انسانها ميباشد.
. همه اعضاي
اين دو گروه
دم را از دست
داده اند. اين
مهم است كه
تذكر داده شود
كه يك صفت
ممكن براي يك
گروه به عنوان
يك صفت اشتقاق
يافته باشد در
حاليكه آن صفت
براي يك گروه
كوچكتر ديگر.به
عنوان مثال
وجود خز براي
تمام tetrapoda يك صفت
اشتقاق يافته
است وبه عنوان
وجه تمايز
آنها از جدشان
يعني خزندگان
ميباشد. اگر
چه آن يك صفت
اوليه براي
همه
پستانداران ميباشدو
براي تعيين
رابطه تكاملي
در پستانداران
مفيد نميباشد.
مثال
ديگر وجود يك
تخم آمنيوتي
به عنوان يك
صفت اشتقاق
يافته براي
آمنيون داران
مي باشد.

فرضيه
اصلي يك
آناليز
كلاديستيك
اين است كه بوسيلهexamining site صفات
اوليه و صفات
اشتقاق يافته
ميتوان
دياگرام آنها
رارسم كرد و
روابط تكاملي بين
گروهها
رامشخص كرد.
نقاط انشعاب
يافته كه در
دياگرام وجود
دارد در هر زمان
به عنوان يك
صفت اشتقاق
يافته است كه
يك گروه داراي
اين صفت ميباشد
و گروه ديگر
آن صفت را
ندارند.
در
مثال زير 5
گروه ((fish-reptile-monkeys-apes-man از
موجودات را در
نظر گرفته و
بر اساس صفات
اوليه و
اشتقاق يافته
كلادوگرام
آنها را رسم
ميكنيم :4 صفت
را در اين 5
گروه مورد
بررسي قرار ميدهيم:
1_وجود اندام
حركتي2 -وجود
خز 3-داشتن دم
4-راه رفتن بر
روي دوپا
ماهي
به عنوان گروه
خارجي كه هيچ
كدام از صفات
را ندارد.
وجود4 اندام
حركتي در 4
گروه ديگر به
عنوان يك صفت
اوليه است
ونشان ميدهد
كه اين 4 گروه
از يك جد
مشترك مي باشند.
صفت ديگر وجود
خز است كه در monkeys, apes, man
وجود داردبه
عنوان يك صفت
اشتقاق يافته
براي اين 3
گروه است
داشتن دم كه
در apes ,man ديده مي
شود و در ديگر
گروهها ديده
نميشود.
وصفت
چهارم صفت راه
رفتن بر روي
دو پا كه فقط
در انسانها
ديده ميشود
كه يك صفت
اشتقاق يافته
است. باعث


جدایی
تولید مثلی
طبق
مفهوم بیولوژیکی
گونه که بوسیله
Ernest Mory در
سال 1940 ارائه شد:
گونه گروهی از
جمعیت هاست که
می توانند با
هم آمیزش کنند
و افراد زنده
ماندنی و
بارور تولید می
کنند. این
گروه از جمعیت
ها نمی توانند
به طور طبیعی
با گونه های دیگر
جفتگیری کنند
که این به دلیل
مکانیسم های
جدایی تولید
مثل است. 2 مکانیسم
جدایی تولید
مثلی وجود
دارد:
Prezygotic و Postzygotic
Prezygotic
قبل از اتحاد
گامت هاست و Postzygotic بعد از
اتحاد گامت
هاست.
یکی
از مکانیسم های
Prezygotic ، Temporal Islation (
جدایی موقتی)
است. این جدایی
از لقاح جلوگیری
می کند، زیرا
دو گونه
متفاوت در
زمان های
مختلف تولید
مثل می کنند.
دوره های زمانی
می توانند به
صورت ساعتی یا
فصلی باشند.
اگر یک گونه
در بهار و
گونه دیگر در
پاییز تولید
مثل کنند قادر
به تولیدمثل
با یکدیگر نیستند.
جدایی
رفتاری ( Behavioral
Isolation) : مکانیسم جدایی
دیگری است. این
مکانیسم از طریق
الگوهای
رفتاری مربوط
به معاشقه عمل
می کند. مثلا
اگر یک گونه،
رفتار معاشقه
ای ویژه ای را
نشان دهد که
از طرف گونه دیگر
تشخیص داده
نشود که این
همان مکانیسمی
است که گرگ ها
را از سگ ها
جدا می کند.
گنجینه ژنی
گرگ و سگ
بصورت متفاوت
حفظ شده است.
گرگ رفتار
تهاجمی تری
نسبت به سگ
دارد، این یک
سد رفتاری ایجاد
می کند که
نتواند رابطه
معاشقه ای
برقرار کند که
این یک مکانیسم
جدایی است.
جدایی
مکانیکی ( Mechanical
Isilation) : به
اندام های تولیدمثل
مربوط می شود.
جدایی مکانیکی
یک سد Prezygotic
است. که از آمیزش
جنسی بین گونه
های متفاوت
جلوگیری می
کند. در این
نوع جدایی می
توان گل را
مثال زد. گلی
که خودلقاح
است، ساختارش
طوری است که
در دسترس
زنبور نیست.
بنابراین این
گیاه به خاطر
مهارهای فیزیکی
که در اندامهای
تولیدمثلی
دارد نمی
تواند با گل
های دیگر هیبرید
ایجاد کند. یا
در damselfly که
پنیس به فرمهای
مختلف باعث می
شود با ماده
های مناسب آمیزش
کنند.

These damselfly penises illustrate just how complex insect
genitalia may be
جدایی
جغرافیایی یا
آلوپاتریک ( Geographial Isolation) :
اصطلاحی است
که برای
مطالعات
تکاملی
استفاده می
شود. وقتی بخشی
از جمعیت یک
گونه از نظر
جغرافیایی از
سایر جمعیت های
آن گونه جدا می
شود، در اثر
گذشت زمان از
نظر خصوصیات
تکاملی از جمعیت
های والد خود
متفاوت می
شوند ( به خاطر
تفاوت ژنتیکی).
اگر
سدهای جغرافیایی
( مثل کوه، رود )
برداشته
شوند، اعضای
دو جمعیت ممکن
است در جفتگیری
با هم نا توان
باشند. بنابراین
جدایی جغرافیایی
یک فاکتور کلیدی
در گونه زایی
است. در واقع
هر چه تفاوتهای
ژنتیکی بیشتر
باشد امکان
مخلوط شدن ژن
ها کمتر و
کمتر می شود.
در نتیجه جدایی
جنسی رخ می
دهد.

Geographic isolation can instigate a speciation event — but genetic changes are necessary to complete the process

جدایی
اکولوژیکی ( Ecological Isolstion) : در این
حالت جمعیت ها
در فضای مشابه
هستند اما در
محل های
متفاوتی زندگی
می کنند. برای
مثال 6 گونه
پشه وجود دارد
که از نظر فیزیکی
مشابه هستند
اما آنها نمی
توانند با هم
تولیدمثل
کنند زیرا در
مکان های
متفاوتی تولیدمثل
می کنند. یکی
در آب راکد و دیگری
در آب روان و ....
مکانیسم
های جدایی
گفته شده را می
توان در آزمایشگاه
از بین برد در
واقع انتخاب
مصنوعی اعمال
کرده و باعث
جفتگیری گونه
های مختلف با
هم شد. اما نمی
توان جلوی جدایی
گامت ها و
سدهای Postzygotic
را گرفت. زیرا
گامت های دو
گونه از نظر شیمیایی
با هم
ناسازگارند
بنابراین از
لقاح جلوگیری
می کنند.
Postzygotic
or Hybrid barriers
اگر
لقاح اتفاق بیافتد
چندین سد برای
هیبرید وجود
دارد.
1.
اولین سد، سد
گامتی است:
گامت ها با
موفقیت با هم
ترکیب می شوند
اما فورا می میرند
قبل از اینکه
تقسیم سلولی
اتفاق بیافتد.
2.
سد دوم، سد
تخمی است که
تخم تشکیل می
شود اما به
سرعت می میرد.
3.
سد سوم، سد جنینی
یا تخمی است
که جنین هیبریدی
بطور خودبه
خودی سقط می
شود.
4.
سد چهارم،
تولید هیبرید
غیر زنده ماندنی
است. که
فرزندان به دنیا
می آیند ولی
مقاوم نیستند
و خیلی زود
تسلیم فشارهای
محیطی می شوند
و می میرند.
5.
سد پنجم، تولید
هیبرید
نابارور است
که زاده ها نمی
توانند تولیدمثل
داشته باشند.
این مکانیسم
ها از چرخش
دوباره مواد
ژنتیکی در
گونه های جدا
از هم جلوگیری
می کند. مثلا mule از آمیزش
اسب ماده و خر
نر بوجود می آید.
اما این هیبرید
نازاست. در
واقع به دلیل
مکانیسم جدایی
Postzygotic این
هیبرید
نازاست و قابلیت
باروری ندارد.
مکانیسم
جدایی prezygotic
و postzygotic
گونه های
مختلف رادر طی
سال حفظ کرده
اند.
تاریخچه
:
تئوری sexual selection ابتدا
توسط داروین
در کتاب The origin of
species
پیشنهاد شد که
البته ابتدا
روی انتخاب طبیعی
متمرکز بود.
داروین در
کتاب بعدی خود
، The descent of man and selection in relation to
sex
، بیشتر در
مورد انتخاب
جنسی صحبت کرد
چراکه احساس
کرد که انتخاب
طبیعی به تنهایی
نمی تواند توجیه
کننده بعضی
سازگاری های غیر
رقابتی باشد
مثل دم در
طاووس نر.
داروین
انتخاب جنسی
را به دو گروه
اولیه تقسیم
کرد : رقابت نر – نر ( که
باعث سازگاری
هایی مثل ایجاد
شاخ در نرها می
شود که در
مشاجره بین
نرها استفاده
می شود ) و
انتخاب ماده (
که باعث
سازگاری هایی
مثل پرهای زینتی
، آوازهای زیبا
و موارد دیگری
که به جلب
توجه و جاذبه
مرتبط است می
شود ).
نظریه
انتخاب جنسی
داروین توسط
والاس به شدت
مورد انتقاد
قرار گرفته است.
او بیان می
کند که رقابت
نر – نر بطور خیلی
ساده بخشی از
انتخاب طبیعی
است.
والاس
همچنین به شدت
با این نظریه
داروین که رنگی
و زیبا بودن
دم طاووس نر
سازش یافته
است ، مخالفت
می کند و بیان
می کند که این
می تواند در
نتیجه تکوین فیزیولوژیکی
باشد.
با وجودی
که داروین
انتخاب جنسی و
طبیعی را دو
فرایند مستقل و
با اهمیت یکسان
می داند اما بیشتر
هم عصران او
متقاعد نشده
اند و بیان می
کنند که
انتخاب جنسی
اهمیت کمتری
نسبت به
انتخاب طبیعی
دارد و می
تواند به
عنوان جزئی از
انتخاب طبیعی
در نظر گرفته
شود.

نظریه
انتخاب جنسی :
نظریه
انتخاب جنسی بیان
می کند که ویژگی
تکاملی خاصی می
تواند توسط
رقابت درون
گونه ای توضیح
داده شود.
داروین
انتخاب جنسی
را به صورت
تأثیرات نزاع
بین افراد یک
جنس ، بطور کلی
نرها ، بر سر
مالکیت جنس دیگر
تعریف می کند.
امروزه زیست
شناسان بین
"نزاع نر با
نر" ، "انتخاب
جفت" ( که بیشتر
ماده آن را
انجام می دهد )
و "تضاد جنسی"
، تفاوت قائل
می شوند. ویژگی
هایی که برای
"نزاع نرها"
انتخاب می شود
صفات ثانویه
جنسی نامیده می
شوند مثل شاخ
ها که گاهی
اوقات هم به
عنوان سلاح در
نظر گرفته می
شوند و صفاتی
که برای
"انتخاب جفت"
انتخاب می
شوند صفات تزئینی
(ornament ) نامیده
می شوند. توجه
زیادی به
انتخاب پنهانی
ماده داده شده
است که پدیده
ای است که در
جانورانی که
لقاح داخلی
دارند مثل
پستانداران و
پرندگان
اتفاق می افتد
و در آن ماده
بدون اطلاع نر
، خود را از اسپرم
های آن رهایی
می بخشدو
معادل آن در "نزاع
نر با نر" ، sperm competition نامیده
می شود. رقابت
مستقیم بین
افراد یک جنس (
معمولاً نرها
) برای جفت ،
همچنین
انتخاب درون
جنسی ( intrasexual selection )
و انتخاب جفت
توسط ماده به
نام انتخاب بین
جنسی ( intersexual selection )
نیز نامیده می
شود. ماده ها
معمولاً ترجیح
می دهند که با
نرهای دارای ویژگی
های تزئینی
جفت شوند. این
ترجیح ماده
برای بعضی
صفات مرفولوژیکی
نر ابتدا توسط
genetic drift افزایش
یافته است که
انتخاب را
متوجه نرهایی
می کند که
صفات تزئینی
بهتری دارند.
این به فرضیه sexy son
معروف است. در
مقابل ژن هایی
که به نر
امکان این را
می دهد که
صفات تزئینی
بهتری نشان
دهد مقاومت بیشتر
در برابر بیماری
ها و متابولیسم
کارآمدی را نیز
ممکن می سازد.
Intrasexual and intersexual selection : در
انتخاب درون
جنسی ، نرها
با صفات تزئینی
خاص ، امتیاز
تولید مثلی
خاصی کسب می
کنند بدون
دخالت ترجیح
فرد ماده. این
امتیاز توسط
سلاح هایی که
در نزاع ها به
کار برده می
شود به دست می
آید. استفاده
از تزئینات
جنسی در فرد
نر ابتدا به
علت ایجاد یک
عدم تقارن در
بین نرهای رقیب
اتفاق می
افتد. صفات
تزئینی جنسی
به عنوان یک
وسیله سیگنال
دهنده در بین
نرها استفاده
می شود تا یک
برتری یابی ایجاد
شود. هنگامی
که دو فرد نر
با یکدیگر درگیری
را آغاز می
کنند این
مواجهه به جایی
خواهد انجامید
که عدم تقارن
ها خود را
توسط استفاده
خشونت بار از
تزئینات جنسی
نشان می دهد.

دو گوزن
نر در حال
نزاع
همچنین
رقابت نر – نر
می تواند شکل
رقابت اسپرمی
به خود بگیرد.
با این
وجود بطور کلی
انتخاب جنسی
به فرایند
انتخاب یک جنس
( معمولاً
نرها ) توسط
جنس مخالف (
معمولاً ماده
ها ) اشاره
دارد. این فرایند
intersexual selection نامیده
می شود. Fisher
اشاره می کند
که این ترجیح
دادن می تواند
کنترل ژنتیکی
داشته باشد و
بنابراین به
ترکیبی از
انتخاب طبیعی
و همچنین تزئینات
جنسی بهتر
اشاره دارد. اینکه در
این نوع
انتخاب کدام
جنس به عنوان
عامل محدود کننده
بیان می شود
توسط Bateman مشخص
شده است. او می
گوید جنسی که
بیشتر برای
تولید مثل
سرمایه گذاری
می کند عامل
محدود کننده
است که جنس
مخالف بر سر آن
به رقابت می
پردازد.
انتخاب
جنسی در انسان
:
چارلز
داروین بیان
کرد که ریش
مردان و همچنین
کم مویی انسان
ها نسبت به
پستانداران
نزدیک آنها می
تواند در نتیجه
انتخاب جنسی
باشد. او بیان
کرد که به دلیل
اینکه بدن
زنان در مقایسه
با مردان موی
کمتری دارد ،
کم مویی یکی
از مواردی است
که در نتیجه
انتخاب مردان
بوجود آمده
است. همچنین
او فرضیه ای
داشت مبنی بر
اینکه نژادهای
مختلف انسانی
به دلیل
انتخاب جنسی
بوجود آمده
اند چراکه
انتخاب طبیعی
نمی تواند
پاسخگوی آن
باشد. George
Miller
که روی رفتار
انسانی بسیار
مطالعه کرده بیان
کرده است که
بسیاری از
رفتارهای
انسانی که در
بقا نقش چندانی
ندارند ، مثل
شوخ طبعی ،
موسیقی ،
هنرهای تجسمی
، تکلم و فعالیت
های بشر
دوستانه ، به
دلیل سازگاری
های در جهت
معاشقه می
باشد. بسیاری
از فرضیه ها
همچنین بیان می
کند که تکامل
مغز بشر نیز یک
ویژگی ایجاد
شده توسط
انتخاب جنسی
است. در این
رابطه می توانیم
از کلمات مثال
بزنیم که بیشتر
انسان ها خیلی
بیشتر از چیزی
که برای
برقراری
ارتباط لازم
است ، می
دانند. Miller
بیان می کند
که این دانش
فراوان به این
علت است که
افراد از
کلمات
استفاده می
کنند تا هوش و
قدرت خود را
به جفت خود
نشان دهند. آزمایشات
نشان می دهند
که مردان
هنگامی که در یک
رابطه عاطفی
قرار دارند
ازکلمات غیر
معمول تر بیشتر
استفاده می
کنند در مقایسه
با زمانی که
در رابطه خاصی
به سر نمی
برند و این
نشان دهنده این
است که کلمات
به عنوان یک
وسیله جنسی
استفاده می
شود.

منبع :
www.wikipedia.org
گونه
زایی : بوجود
آمدن بیش از یک
گونه جدید در
جایی که فقط یکی
از آن گونه
وجود داشته
است .
مهمترین
اصل در گونه
زایی جدایی
تولید مثلی
است که این به
دو طریق امکان
پذیر است : جدایی
پیش زیگوتی و
جدایی پس زیگوتی
مکانیسم
های جدایی پیش
زیگوتی :
جدایی
پیش زیگوتی به
چند طریق
انجام می شود :
جدایی
جغرافیایی ،
ممانعت از ترکیب
گامت ها ، جدایی
مکانیکی ، جدایی
رفتاری ، جدایی
زمانی

در اینجا
به بحث در
مورد جدایی
زمانی می
پردازیم .

در
جدایی زمانی
ازدواج و
تبادل گامتی
در زمانهای
متفاوتی از
سال اتفاق می
افتد. جدایی
زمانی جریان
ژنی بین جمعیت
ها را کاهش می
دهد.این مکانیسم
حتی از تبادل
ژنی بین دو
گونه که با هم
در یک جا زندگی
می کنند جلوگیری
می کند . به طور
مثال در آمریکای
شمالی راسو ها
با هم زندگی می
کنند اما گونه
S.putorius در آخر
زمستان و گونه
S.gracilis در
آخر تابستان mating دارند.

بعضی
از گونه های
حشرات mating را
فقط در چند
ساعت روز یا
شب به صورت
متفاوت انجام
می دهند.
تفاوت در تنظیم
زمانی breeding رایج
است.
مثال:
Photinus fireflier در
زمان های
متفاوتی از
روز دیده می
شود و در جدایی
زمانی تولیدمثل
شرکت دارد.
دردر
دو گونه جیرجیرک
در آمریکای
شمالی گونهGryllus
pennsylvanicus و گونه
G.veletis زمان تولید
مثلی جدا در
پاییز و بهار
دارند.
دو
گونه حشره به
نام های Chrysopa downesi وChrysopa carnea که
هر کدام به
ترتیب در جنگل
های کاج و
جنگل درختان
برگ ریز زندگی
می کنند و طبق
نمودار پایین
زمان تولید
مثل آنها
متفاوت است و
همین باعث جدایی
گونه ای در
آنها شده است.
بعضی
از گونه های
قورباغه نیز
در یک محل
زندگی می کنند
ولی پیک های
فعالیت تولیدمثلی
جدا از هم
دارند. مثال دیگر دو
گونه کاج است
که با هم در
کالیفرنیا
رشد می کنند
گونه radiate Pinus
در فوریه و
گونه P.muricataدر آپریل
گرده افشانی می
کنند و به
خاطر این جدایی
زمانی تولیدمثلی
به دو گونه
جدا تبدیل شده
اند.

آناتومی
مقایسه ای,
وجود ساختارهایی
به نام
ساختارهای
وستیجیال را
آشکار می
سازد. بسیاری
از جانداران
دارای اندام
ها یا بخش هایی
از یک اندام
هستند که به
نظر می رسد
فاقد وظیفه
مشخصی بوده و
تحلیل رفته
اند. این
اندام ها اغلب
کوچکتر از حد
معمول بوده و
فاقد اجزای
اصلی هستند.
ساختارهای
وستیجیال
آثاری به جای
مانده از
ساختارهایی
تکامل یافته
تر هستند که
قبلا در
جانوران
اجدادی وجود
داشته و دارای
وظیفه مشخصی نیز
بوده اند. وجود
اندامهاي
همتا در
جانداران،
نشاندهندهي
وجود رابطهي
تكاملي ميان
آنها و داشتن
منشا مشترك
است .وقتي
آناتومي جانداران
مختلف را
مقايسه ميكنيم،
به
ساختارهايي
بر ميخوريم
كه در يك عده
بسيار كارآمد
هستند اما در
عدهي ديگر،
كوچك شده و
حتي ممكن است
فاقد كار خاصي
باشند. مثلا
بيشتر
پرندگان، بالهايي
كارآمد دارند
كه بهكمك آن
پرواز ميكنند
اما در
شترمرغ، اين
بالها آنچنان
كوچك و ضعيف
شدهاند كه
پرواز با آنها
ممكن نيست.
اين اندامهاي
كوچك و ضعيف
شده را اندامهاي
وستيجيال (به
معني ردپا) ميناميم.
در واقع اندام
وستيجيال
ردپاي تغيير گونهها
هستند. مارها
با اينكه پا
ندارند اما
بقاياي پا در
لگن آنها بهصورت
وستيجيال
موجود است و
اين حاكي از
وجود رابطهي
مشترك تكاملي
ميان مارها و
ديگر مهرهداران
است. در
بدن انسان بیشتر
از 100 ساختار
وستیجیال
وجود دارد,
مانند
استخوان
دنبالچه, سومین
دندان آسیا
(دندان عقل) و
ماهیچه هایی
که گوش هایمان
را حرکت می
دهند. خوک ها
دارای پنجه های
وستیجیال
هستند, پرندگان
بدون بالی مثل
کیوی (نوعی
مرغ نیوزلندی)
دارای
استخوان های
بال وستیجیال
هستند و بسیاری
از جانوران
کوری که در
لانه های زیر
زمینی یا
غارها زندگی میکنند,
دارای چشم های
وستیجیال غیر
فعال می
باشند. تشکیل
ساختار وستیجیال
در ارتباط با
وقوع سازش در یک
گونه , در
هنگام تغییر
شرایط محیط
زندگی می
باشد. در صورت
پدید آمدن چنین
شرایطی, بعضی
از ساختار ها
اهمیت کمتری
در حفظ بقا پیدا
کرده وممکن
است در جریان
تکامل فقط اثری
در آنها باقی
بماند. وقتی یک
ساختار
بزرگتر , برتری
را به جاندار
اعطا نکند,
ممکن است در
طول زمان کوچک
تر شده و همه یا
بخش اعظم وظایفش
را از دست
بدهد. وجود
ساختارهای
وستیجیال
اغلب ضرری برای
جاندار ندارد,
از این رو
انتخاب طبیعی
قادر به حذف
کامل آن نبوده
و ساختار وستیجیال
در بسیاری از
نسل های بعدی
حفظ می شود.
نواحی
هیبریدی بیش
از70سال
هست که مورد
تحقیق قرار
گرفتند.در سال
1989بارتون
وهویت
صدوهفتاد ناحیه
ی هیبریدی را
گزارش
دادندوبیان
داشتند که
نواحی هیبریدی
یک ناحیه ی
جغرافیایی
است که دو جمعیت
واگرا دریک حد
جغرافیایی
باهم برخورد میکنند
ودر ضمن آمیزش
زاده های هیبریدی
تولید میکنند
که این هیبریدها
درآن جا شیوع یافته
وتشکیل شیب (cline) می
دهند.

یکclineیک شیب
جغرافیایی با
فرکانسهای یک
ژن میباشدوانواع
مختلفی دارد
مثلا نواحی هیبریدی
جز شیبهای پله
دار میباشند
که این نوع از cline هادارای
شیب های چشمگیری
از ژنها میباشند. یک ناحیه ی هیبریدی
توسط مراحل
تقویت کننده میتواندبه
درون یک سد
گونه ای گسترش
یابد.هم چنین
انواع دیگرگونه
های موجود در
این نواحی با
گونه های هیبریدزا
تفاوتهای
اساسی دارند
نمونه ی کلاسیک
از این گونه
ها گاوهای carrionوhoodedمی
باشند که باهم
آمیزش کرده
وتولید هیبریدهایی
در طول یک
محدوده در
اروپای مرکزی
میکنند.
نواحی
هیبریدی به دو
دسته تقسیم میشوند:
1.نواحی
هیبریدی اولیه
2.نواحی
هیبریدی ثانویه
خصوصیات
نواحی هیبریدی
اولیه :
این
نواحی محلهای
جغرافیایی
هستندکه گونه
های موجود
درآن دارای
تفاوت درون
خود بوده
وشکلهای هیبریدیشان
باهم ارتباط
وآمیزش
دارند.مثل
گونه زایی
پاراپاتریک
خصوصیات
نواحی هیبریدی
ثانویه:
در این
نواحی ادامه ی
جمعیت انتشار یافته
بوسیله ی یک
سد جداکننده
تقسیم میشود.
مثل
نواحی جغرافیاییrefugiaیا جمعیتهای
جداشده ی
جغرافیایی
قابل تشخیص در
گونه زایی
آلوپاتریک

پس
وجود نواحی هیبریدی
یک مرحله ی
اساسی است برای
اینکه گونه زایی
پاراپاتریک
وآلوپاتریک
اتفاق بیافتد.
یک
شکل ازنتایج
نواحی هیبریدی
جایی است که یک
گونه در آن
متحمل گونه زایی
آلوپاتریک میشود
به طوری که دو
جمعیت جدید
بعد از یک
دوره ی جدایی
جغرافیایی با
هم جفت گیری
کرده و ایجاد
زاده های هیبریدی
میکنند که
دارای ترکیبی
از آللهای
مشخص هر دو
جمعیت میباشند
هم چنین جریان
ژنی جدیدی هم
وارداین ناحیه
ی هیبریدی میشود. این
ژنها در بین
جمعیتها بوسیله
ی آمیزش میان
افراد هیبرید
وغیر هیبرید میتوانند
در جریان
وگردش باشند. درنهایت
این مراحل
وپروسه ها در
گونه زایی
آلوپاتریک
درنهایت
منجربه شکل گیری
یک )شیب)clineمیان
دو جمعیت خالص
در نواحی هیبریدی
میشود . در این نواحی
هیبریدی در بسیاری
از حالات زاده
های هیبریدی
شایستگی کمتری
نسبت به خویشاوندان
خود دارند چرا
که فاقد
کمپلکس های ژنی
کامل موجود در
خویشاوندان
خودمی باشند. نواحی
هیبریدی هم چنین
در نواحی که
در آنها گونه
زایی
پاراپاتریک
اتفاق می افتد
دیده میشود در
واقع
پاراپاتری
زمانی است که
دو گونه یا زیرگونه
بدون سد جغرافیایی
در کنارهم
زندگی کنند
بدون اینکه
باهم اختلاط یابند
ولی بعدابطور
ثانویه به هم
بپیوندند
واختلاط در
مرزهای وسطی
که در این مرز
دوگونه باهم
اورلپ دارند
صورت گیردودر
نهایت با تولید
زاده های هیبریدی
تولید ناحیه ی
هیبریدی کنند.
توضیحات
همراه با شکل
نواحی هیبریدی
اولیه وثانویه:
نواحی
هیبریدی ثانویه:
نواحی
هیبریدی ثانویه
شیبهای خنثی
هستندوژنهای
انتخابی این شیبها
کاملا بهم
وابسته بوده
ودرنقطه ی
شروع هیبریداسیون
قرار
دارندالبته
قبل ازاینکه
تفاوتهای ژنتیکی
درجدایی
اتفاق بیافتد.

درنواحی
هیبریدی اولیه
فقط ژنهای
پاسخ دهنده به
انتخاب های
خاص
وجوددارند که
گونه هایی در
مکانهای اولیه
تولیدمیکنندکه
درنقطه شروع هیبریداسیون
قرارمیگیرند.

ژنهای
طبیعی –هیبریداسیون-نواحی
هیبریدی
درپنج
حالت میتوان
ارتباط اینها
را بیان کرد:
1.درحالت
هیبرید زون
همه ی ژنها در
تفاوتهای
زاده ها در
نواحی جغرافیایی
مختلف درگیر نیستند.
2.پدیده
ی نوترکیبی در
هیبریدها بوسیله
ی ژنهای جدا
شده از ژنهای
طبیعی میتواند
صورت گیرد.
3.ژنهای
تحت انتخاب یک
انتشار بین cline ای
درآللهاشان
خواهندداشت.

4.ژنهای
طبیعی ممکن
است بوسیله ی
تعدادی از
گونه ها انشار
یابند.

5.ژنهای
مختلف پاسخ
دهنده ی به
انتخاب های
مختلف میتوانند
clineهای
مختلف نشان
دهند.

Synapomorphy &
Symplesiomorphies
Not
downloaded
Heterochrony
PDF
Do not downloaded
تکامل
اسبها
اسبها
یکی از تاریخی
ترین جانوران
اهلی زنده که
در زندگی
انسانها یک
نقش حیاتی به
مدت دهها هزار
سال گذشته بازی
کرده است. هیپچ
جانور دیگر،
اهلی یا وحشی،
همانند اسب چنین
اثر بزرگی روی
تمدن نداشته
اند. چنانچه
در جنگها، حمل
و نقل،
مسافرت، هنر و
ورزش از این
جانور
استفاده کرده
اند.
تاریخچه:
در
سال 1870
دیرین شناسی
بنام Marsh یک تشریح
کاملی از فسیل
اسبهای کشف
شده آمریکای
شمالی را
منتشر کرد. سری
فسیلهای اسب
که Marsh
نشان داده
بود، یک مثال
مهمی از تکاملی
که به نسل
واحدی منتهی می
شد، بود.
چندین
سال بعد، موزه
آمریکایی Natural History
که یک مجموعه
کاملی از فسیل
ها را جمع
کرده بود که
برای نشان
دادن تکامل
تدریجی اسبها
از Hyracotherium تا Equus
کنونی بود. پس
فسیل اسبها به
خاطر اینکه از
55 میلیون
سال پیش کشف
شده و تکامل
تدریجی را از Hyracotherium
تا Equus
نشان می دهد
دارای اهمیت زیادی
است، علی رغم
اینکه تمام فسیل
اسبها کشف
نشده اند، برای
تکاملیون و
تاکسونومیستها
مهم می باشد.
مشخصات
رده بندی:
|
Kingdom: Animally Phylum: Chordate Class: Mammalian Order: Perissodactyla Family: Equidae Genus: Equus Species: E. caballus |

درخت
تکاملی اسبها
و دوره های زمین
شناسی:
Ø
10000
سال پیش تا
حالا (اخیر)
Ø
Pleistocene از .01 تا 2.5 میلیون
سال پیش
Ø
Pliocene از 5.3 تا 2.5 میلیون
سال پیش
Ø
Miocene از 24 تا 5.3 میلیون
سال پیش
Ø
Oligocene
از 34
تا 24
میلیون سال پیش
Ø
Eocene از 55 تا 34 میلیون
سال پیش
نظریه
های تکاملی:
1.
تکامل
بصورت خط مستقیم:
همانطوریکه
فسیل های جمع
آوری شده توسط
Marsh نشان می
دادند، گویا
اسبها یکی پس
از دیگری
بوجود آمده و
سپس منقرض شده
اند و جای خود
را به دیگری
داده، تا اینکه
به اسبهای
امروزی رسیده
اند.
2.
تکامل
بصورت درخت پر
شاخ و برگ: این
نظریه بیان می
دارد که در
دوره هایی خاص
اشتقاق گرفته
و شاخه شاخه
شده اند ولی
در نهایت یکی
از شاخه ها این
راه را ادامه
داده و بقیه
آنها منقرض
شده اند.
همانند اینکه
شاخه های درختی
را قطع کنیم و
فقط تنه آن
باقی ماند.
امروزه نیز این
نظریه مورد
قبول است.
اسبهای
دوره Eocene:
این
اسبها شامل Hyracotherium،
Eohippus، Epihippus
بودند. همگی
جز جانوران
جنگلی بودند و
شبیه هیچ یک
از اسبهای
کنونی نبودند.
اسبهای این
دوره تفاوت زیادی
با سایر تک سم
داران مانند
کرگدنها و
خوکها
نداشتند، بر
عکس اسبهای
امروزی که با
سایر جنسهای
خانواده خود
تفاوتهایی را
نشان می دهند.
مشخصات
Hyracotherium:
1.
ظاهری
سگ مانند، پشت
کمان مانند،
گردن کوتاه
2.
پوزه
کوتاه، دم طویل
3.
تغذیه
از میوه جات و
شاخ و برگ (همه
چیز خوار)
4.
مانند
گوزن کوچک آسیایی
چهار نعل و
آهسته راه می
رود.
5.
پاهای
قابل انعطاف و
قابل چرخش با
استخوانهای
اصلی
6.
چهار
انگشت در پای
جلویی و سه
انگشت در پای
عقبی
7.
دندانهایی
با تاج کوتاه،
3
دندان آسیاب
اصلی، 4
دندان آسیاب
کوچک و یک
دندان گرگی
(دندانهای همه
چیزخوار)

اسبهای
Oligocene :
همان
اسبهای متوسط
اندازه می
باشند که
شامل: Eohippus، Miohippus
می باشند. در
اثر تغییرات
آب و هوایی که
منجر به خشک
تر شدن هوا
شده است، رفته
رفته جنگلهای
وسیع جای خود
را به
علفزارها و
چمنزارها
داده اند. در اثر
این تغییرات،
تغییراتی در
دندانها و
تاجهای اسبهای
این دوره به
وقوع پیوست
چنانچه
دندانها سفت و
بزرگتر شدند.
Mesohippus:
گونه
های جدید در این
دوره (40
میلیون سال پیش)
بوجود آمدند.
1.
این
اسبها اندکی
از اسبهای ما
قبل خود
بزرگتر بودند.
اسبهای این
دوره دیگر
ظاهری سگ مانند
نداشتند.
2.
پشت
فاقد حالت
کمانی، پاهای
بزرگتر، گردن
درازتر، صورت
کشیده تر
3.
یک
حفره در صورت
و یک گود شدگی
در جمجمه
4.
روی
پاهای جلویی و
عقبی 3
انگشت دارد و
انگشت چهارم
فقط بصورت اثری
از آن باقی
مانده است.


Miohippus (36
میلیون سال پیش
بوجود آمدند):
1.
یک
نمونه میوهیپوس
بطور قابل
توجهی از مزوهیپوس
بزرگتر بوده و
همچنین دارای یک
جمجمه کشیده
تر بوده است.
2.
حفره
های صورت بیشتر
و عمیق تر
گسترش یافته
است.
3.
گونه
های میوهیپوس
با ظهور یک
تاج بزرگ متغیر
روی دندانهای
فوقانی شروع می
شوند.
این نمونه
مثال خوبی از
اینکه یک صفت
چگونه می
تواند باعث
تنوع در جمعیت
نیاکان خود
شود.
.
اشتقاق میوهیپوس:
حدود 24
میلیون سال پیش
در میوسن اولیه،
میوهیپوس
شروع به گونه
زایی کرد. Equudae
شروع به
انشعاب در دو
خط تکاملی اصلی
و یک انشعاب
فرعی کوچک
کرد:
1.
چراکننده
های 3
انگشتی که anchiteres
نامیده می
شوند، خیلی
موفق بودند و
در سطح زمین
باستان به خوبی
پخش شدند و به
مدت 10 میلیون
سال پیشروی
کردند.
2.
اسبهایی
که برای بدست
آوردن مزایای
گیاهان جدید
از حالت
چراکننده همه
چیزخوار(browsing)
به چراکننده
های گیاهی (grazing)
تغییر کرده
بودند.
3.
یک
انشعاب فرعی
شامل اسبهای
کوتاه بعنوان
مثال Archeohippus
بودند. اینها
خیلی زنده
نماندند.
اسبهایی با
دندانهای تاج
بلند و پاهای
ارتجاعی (18 میلیون
سال پیش):
1.
در این
اسبها
دندانها برای
جویدن علف های
خشن و ساینده
تغییرات
مناسبی را یافته
بودند.
2.
این
اسبها برای دویدن
اختصاص یافته
بودند. همچنین
اندازه بدن،
طول پاها و
درازای صورت
آنها بیشتر
شده بود.
3.
این
اسبها به جای
سگ مانند راه
رفتن شروع به
ایستادن روی
پنجه ها کرده
بودند که این
نیز سازش دیگری
برای افزایش
سرعت آنها
بوده است.
اسبهای
Miocene :
این اسبها
حدود 18
میلیون سال پیش
می زیسته اند
و شامل Kalobatippus ،Parahippus وMerychippus
بودند.
Merychippus:
1.
پوزه
کشیده،
آرواره های عمیق،
چشمها به دور
از پشت جانور
جابه جا شده
است. این تغییرات
از اسب مری چیپوس
یک اسب زیرک،
باهوش نسبت به
سایر اسبهای
اولیه ساخته بود.
2.
مری
چیپوس هنوز 3
انگشتی بود
اما دارای
پاهای کاملاً
ارتجاعی بود.(spring-foot )
3.
زند
زبرین و زند زیرین
به همدیگر ترکیب
شده بودند
آنچنانچه
پاها دیگر
چرخش نداشتند.
تمام این تغییرات
پاهای مری چیپوس
را فقط برای یک
عمل ممکن
ساخته بود: سریع
دویدن در زمینهای
سخت.

اشتقاق
مری چیپوس:
این
اسبها جز اولین
اسبهای چابک
واجد شرایط
بود و به سرعت
تحت تاثیر
گونه زایی
قرار گرفت و
به 19
گونه اسب
چراکننده از
علفزارها در
سه گروه اصلی
تغییر یافت.
که به این
انفجار بزرگ
تکاملی،
اشتقاق Merychippine
می گویند.
1.
چرنده
های سه انگشتی
که بعنوان hipparions
نامیده می
شوند.
2.
یک
خط انشعابی از
اسبهای کوچک
شامل Protohippus وCalippus که
در مجموع
پروتو هیپین
نامیده می
شوند.
3.
اسبهای
حقیقی
سراسر
تکامل اسبها
به نسلهای مری
چیپین وابسته
هستند. حفره
های صورت آنها
عمیق تر شده و
تغییرات آنها
خیلی
استادانه می
باشد.
اسبهای
تک انگشتی
دوره Pliocene:
این
اسبها شامل Polihippus
، Dinohippus و Astrohippus
بودند.
Dinohippus:
سرانجام
اسبهای تک
انگشتی گروه
سوم بنام Dinohippus
ها در حدود 12 میلیون
سال پیش بوجود
آمدند. نیاکان
اصلی آنها
هنوز هم مشخص
نیستند.این
اسبها بطور
شگفت انگیزی
از نظر
موفولوژی پا،
دندان و جمجمه
شبیه Equus ها
بودند.
دندانهای
آنها از
دندانهای مری
چیپوس راست تر
و حفره صورت
آنها کاهش یافته
بود.
در
پایان دوره پلیوسن،
Dinohippus ها یک
کاهش تدریجی
در حفره صورت،
راست شدن
دندانها و دیگر
تغییرات را
نشان دادند.
Equus ها:
سرانجام
حدود 4
میلیون سال پیش
Equus ها بوجود
آمدند که دارای
مشخصات زیر
بودند:
1.
اولین
اسب Equus
اندازه کوچکی
نسبت به اسب
امروزی داشت.
2.
پشت
بدن خیلی سفت،
گردنی دراز و
پاهایی طویل
داشت.
3.
استخوانهای
پا جوش خورده
و بدون چرخش
بودند.
4.
دماغ
کشیده، پوزه
قابل انعطاف و
آرواره عمیق
تر داشتند.
5.
دندانهای
چراکننده
راست با تاج
پوشیده توسط
لایه سیمان.
در طول اولین
دوره یخبندان
در پلیوسن اخیر(2.6 میلیون
سال پیش) گونه
های Equus با
جهانی باستانی
روبرو شدند.
بعضی ها به
آفریقا
مهاجرت کرده و
بصورت
گورخرهای
امروزی تنوع یافتند.
بعضی دیگر در
آسیا و اروپا
بعنوان اسبهای
حقیقی منتشر یافتند
و بعضی نیز در
آمرکای جنوبی
پخش شدند. جنس Equus
شاید تنها
جانور تک سم
موفق به زیستن،
قبل از اینکه
توسط انسان
رام شود، بود.
اسبهای کنونی:
اگرچه اسبهای
تک انگشتی خیلی
موفق بودند
اما Equus ها
یک میلیون سال
پیش در تمام
آفرقا، آسیا،
اروپا، آمریکای
شمالی و آمریکای
جنوبی گسترش یافتند.(بصورت
مهاجرتهایی
با گله های
بزرگ) در دوره Pleistocene یک
انقراض وسیعی
روی
پستانداران
آمریکای شمالی
و جنوبی رخ
داد و منجر به
کشته شدن تمام
اسبهای قاره
آمریکا شد.
برای اولین
بار در دهها میلیون
سال در آمریکا
هیچ اسبی وجود
نداشت. که
بعدها توسط
مهاجرین
اروپایی به
آنجا انتقال یافت.
گونه هایی از
خانواده Equidae که
تا به امروز
زنده مانده
اند شامل:
1.
E. zebra : گورخر
کوهستانی آفریقای
جنوبی
2.
E. caballus: اسبهای
حقیقی که دارای
چندین زیرگونه
هستند.
3.
E. hemi onus : گورخر
کوچک سازش یافته
با بیابانهای
آسیا
4.
E. sinus : الاغ های
حقیقی


| N | First Name | Last Name | Subject |
| 1 | فاطمه | ابوطالبی اردکانی | Natural Selection |
| 2 | فاطمه | اشرفی سلطان احمدی | exaptation |
| 3 | مینا سادات | امامیان | Mutation |
| 4 | میترا | امیری خبوشان | Genetic Drift |
| 5 | فاطمه | تاجبخش | Reproductive Isolation |
| 6 | سحر | جوکار | Habitate Isolation |
| 7 | سکینه | حسین نژاد | Temporal Isolation |
| 8 | مهرنوش | خدائیان | Sexual Selection |
| 9 | عرفان | خسروی | heterochrony |
| 10 | روشنک | راشدی | zone hybride |
| 11 | الهام | رضا زاده | Mechnical Isolation |
| 12 | زهرا | سالارفر | Gametic Isolation |
| 13 | مژگان | سرحدی | Postzygotic Isolation |
| 14 | طیبه | صالحی | Cladistics |
| 15 | مهدیه | عاشق آبادی | Phenetics |
| 16 | حسن | عباسی | Speciation Dynamics |
| 17 | شهریار | علی پور | eqause evolution |
| 18 | فاطمه | فیروزی | primitive&drived characters |
| 19 | سارا | قرباتی | Synapomorphy & Symplesiomorphies |
| 20 | خدیجه | کاظمی | Sym, Allo & parapatry |
| 21 | سمیه | کشتکار | polyploidy |
| 22 | سیما | کلانتری | Adaptive Radiation |
| 23 | مهسا سادات | کمونه | allometric growth |
| 24 | امیر | مجتبوی | Vestigial structure & speciation |
| 25 | فاطمه | مجیدی | Homology & Analogy (Morpho&molecular) |
| 26 | فاطمه | محققی | Behavioral Isolation |
| 27 | منصوره | محمدی | Convergent,Divergent,Parallel & coevolotion |
| 28 | روشنک | مسلمی | Founder Effect, botlleneck in speciation |
| 29 | معصومه | معتمدی | parapathry |
| 30 | طاهره | میرجلیلی | Homeotic genes in Speciation |
| 31 | معصومه | ندرلو | Geographic Isolation |
| 32 | فرشته | نصری | Parsimony &maximum liklihood |
| 33 | مریم | وفاجو | Competition |
| 34 | فاطمه | همتی | fossil dating |